leerdoelstellingen
- Beschrijf de bindweefsellagen verpakking skeletspieren
- leg uit hoe spieren met pezen werken om het lichaam te bewegen
- Identificeer gebieden van de skeletspiervezels
- beschrijven excitatie-contractie koppeling
het bekendste kenmerk van het skelet spier is het vermogen om samen te trekken en beweging te veroorzaken. Skeletspieren werken niet alleen om beweging te produceren, maar ook om beweging te stoppen, zoals weerstand bieden aan de zwaartekracht om de houding te behouden. Kleine, constante aanpassingen van de skeletspieren zijn nodig om een lichaam rechtop of in evenwicht te houden in elke positie. Spieren voorkomen ook overmatige beweging van de botten en gewrichten, het behoud van de stabiliteit van het skelet en het voorkomen van beschadiging of vervorming van de skeletstructuur. Gewrichten kunnen worden verkeerd uitgelijnd of volledig ontwricht door te trekken op de bijbehorende botten; spieren werken om gewrichten stabiel te houden. Skeletspieren bevinden zich door het hele lichaam bij de openingen van interne traktaten om de beweging van verschillende stoffen te controleren. Deze spieren kunnen functies, zoals slikken, urineren en ontlasting, onder vrijwillige controle. Skeletspieren beschermen ook interne organen (met name buik-en bekkenorganen) door als externe barrière of schild voor externe trauma ‘ s te fungeren en door het gewicht van de organen te ondersteunen.
skeletspieren dragen bij tot het behoud van homeostase in het lichaam door warmte te genereren. Spiercontractie vereist energie, en wanneer ATP wordt afgebroken, warmte wordt geproduceerd. Deze warmte is zeer merkbaar tijdens het sporten, wanneer aanhoudende spierbewegingen de lichaamstemperatuur doen stijgen, en in gevallen van extreme kou, wanneer rillen willekeurige skeletspiercontracties veroorzaakt om warmte te genereren.

figuur 1. De Drie Bindweefsellagen. Bundels van spiervezels, genaamd fascicles, worden bedekt door de perimysium. Spiervezels worden bedekt door het endomysium.
elke skeletspier is een orgaan dat bestaat uit verschillende geïntegreerde weefsels. Deze weefsels omvatten de skeletspiervezels, bloedvaten, zenuwvezels, en bindweefsel. Elke skeletspier heeft drie lagen bindweefsel (genaamd “mysia”) die het omsluiten en structuur bieden aan de spier als geheel, en ook compartimenteren de spiervezels in de spier (figuur 1). Elke spier is verpakt in een schede van dichte, onregelmatige bindweefsel genoemd epimysium, die een spier laat samentrekken en krachtig bewegen met behoud van zijn structurele integriteit. Epimysium scheidt ook spier van andere weefsels en organen in het gebied, toestaand de spier om zich onafhankelijk te bewegen.
binnen elke skeletspier zijn spiervezels georganiseerd in individuele bundels, elk een fascicle genoemd, door een middelste laag bindweefsel genaamd de perimysium. Deze fascicular organisatie is gemeenschappelijk in spieren van de ledematen; het staat het zenuwstelsel toe om een specifieke beweging van een spier te activeren door een subset van spiervezels binnen een bundel, of fascicle van de spier te activeren. Binnen elke fascicle, elke spiervezel is ingekapseld in een dunne bindweefsel laag van collageen en reticulaire vezels genaamd de endomysium. Het endomysium bevat de extracellulaire vloeistof en voedingsstoffen ter ondersteuning van de spiervezel. Deze voedingsstoffen worden via het bloed aan het spierweefsel geleverd.
in skeletspieren die met pezen werken om botten aan te trekken, verstrengelt het collageen in de drie weefsellagen (de mysia) met het collageen van een pees. Aan de andere kant van de pees, het fuseert met het periosteum coating van het bot. De spanning die door samentrekking van de spiervezels wordt dan overgebracht door de mysia, naar de pees, en vervolgens naar het periosteum te trekken op het bot voor de beweging van het skelet. Op andere plaatsen, kan de mysia samensmelten met een brede, peesachtige plaat genaamd een aponeurose, of aan fascia, het bindweefsel tussen huid en botten. Het brede bindweefsel in de onderrug waar de latissimus dorsi spieren (de “lats”) in versmelten is een voorbeeld van een aponeurose.
elke skeletspier wordt ook rijkelijk gevoed door bloedvaten voor voeding, zuurstofafgifte en afvalverwijdering. Bovendien wordt elke spiervezel in een skeletspier geleverd door de Axon-tak van een somatisch motorneuron, die de vezel signaleert om samen te trekken. In tegenstelling tot cardiale en gladde spieren, de enige manier om functioneel contract een skeletspier is door het signaleren van het zenuwstelsel.
Skeletspiervezels
omdat skeletspiercellen lang en cilindrisch zijn, worden ze gewoonlijk spiervezels genoemd. Skeletspiervezels kunnen vrij groot zijn voor menselijke cellen, met diameters tot 100 µm en lengtes tot 30 cm (11,8 in) in de sartorius van het bovenbeen. Tijdens de vroege ontwikkeling fuseren embryonale myoblasten, elk met zijn eigen kern, met tot honderden andere myoblasten om de meerkernige skeletspiervezels te vormen. Meervoudige kernen betekenen meerdere kopieën van genen, waardoor de productie van de grote hoeveelheden eiwitten en enzymen die nodig zijn voor spiercontractie.
sommige andere terminologie geassocieerd met spiervezels is geworteld in het Griekse sarco, wat “vlees” betekent.”Het plasmamembraan van spiervezels wordt de sarcolemma genoemd, het cytoplasma wordt sarcoplasma genoemd en het gespecialiseerde gladde endoplasmatische reticulum, dat calciumionen opslaat, loslaat en ophaalt (Ca++) wordt het sarcoplasmatische reticulum (Sr) genoemd (Figuur 2). Zoals binnenkort zal worden beschreven, is de functionele eenheid van een skeletspiervezel het sarcomeer, een sterk georganiseerde opstelling van de contractiele myofilamenten actine (dunne gloeidraad) en myosine (Dikke gloeidraad), samen met andere ondersteunende eiwitten.

Figuur 2. spiervezel. Een skeletspiervezel is omgeven door een plasmamembraan genaamd de sarcolemma, die sarcoplasma, het cytoplasma van spiercellen bevat. Een spiervezel bestaat uit vele fibrillen, die de cel zijn gestreepte uiterlijk geven.
het sarcomeer
het dwarsgestreepte uiterlijk van skeletspiervezels is te wijten aan de opeenvolging van de myofilamenten van actine en myosine van het ene uiteinde van de spiervezel naar het andere uiteinde. Elk pakket van deze microfilamenten en hun regulerende eiwitten, troponine en tropomyosine (samen met andere eiwitten) wordt een sarcomeer genoemd.
het sarcomeer is de functionele eenheid van de spiervezel. Het sarcomeer zelf is gebundeld in de myofibril die de gehele lengte van de spiervezel loopt en zich aan het sarcolemma aan het einde ervan hecht. Als myofibrils samentrekken, trekt de hele spiercel samen. Omdat myofibrillen slechts ongeveer 1,2 µm in diameter hebben, zijn er honderden tot duizenden (elk met duizenden sarcomeren) te vinden in één spiervezel. Elk sarcomeer is ongeveer 2 µm lang met een driedimensionale cilinder-achtige opstelling en wordt begrensd door structuren genaamd z-schijven (ook Z-lijnen genoemd, omdat foto ‘ s zijn tweedimensionaal), waaraan de actin myofilamenten zijn verankerd (Figuur 3). Omdat het actine en zijn troponine-tropomyosine complex (projecteren van de Z-schijven naar het centrum van het sarcomeer) vormen strengen die dunner zijn dan de myosine, wordt het de dunne gloeidraad van het sarcomeer genoemd. Evenzo, omdat de myosine strengen en hun meerdere hoofden (projecteren vanuit het centrum van de sarcomere, naar maar niet helemaal naar, de Z-schijven) hebben meer massa en zijn dikker, ze worden genoemd de dikke gloeidraad van de sarcomere.

Figuur 3. De Sarcomere. De sarcomere, het gebied van de ene Z-lijn naar de volgende Z-lijn, is de functionele eenheid van een skeletspiervezel.
de neuromusculaire junctie
een andere specialisatie van de skeletspieren is de plaats waar de terminal van een motorneuron de spiervezel ontmoet-de neuromusculaire junctie (NMJ) genoemd. Hier reageert de spiervezel voor het eerst op het signaleren door het motorneuron. Elke skeletspiervezel in elke skeletspier wordt geïnnerveerd door een motorneuron bij de NMJ. Excitatiesignalen van het neuron zijn de enige manier om de vezel functioneel te activeren om samen te trekken.
excitatie-contractie koppeling
alle levende cellen hebben membraanpotentialen of elektrische gradiënten over hun membranen. De binnenkant van het membraan is gewoonlijk rond -60 aan -90 mV, met betrekking tot de buitenkant. Dit wordt bedoeld als het membraanpotentieel van een cel. Neuronen en spiercellen kunnen hun membraanpotentialen gebruiken om elektrische signalen te genereren. Ze doen dit door de beweging van geladen deeltjes, ionen genaamd, over hun membranen te regelen om elektrische stromen te creëren. Dit wordt bereikt door gespecialiseerde proteã nen in het membraan genoemd ionenkanalen te openen en te sluiten. Hoewel de stromen die door ionen worden geproduceerd die zich door deze kanaalproteã nen bewegen zeer klein zijn, vormen zij de basis van zowel neurale het signaleren als spiercontractie.
zowel neuronen als skeletspiercellen zijn elektrisch prikkelbaar, wat betekent dat ze actiepotentialen kunnen genereren. Een actiepotentiaal is een speciaal type elektrisch signaal dat langs een celmembraan als golf kan reizen. Hierdoor kan een signaal snel en getrouw over lange afstanden worden verzonden.
hoewel de term excitatie-contractie koppeling sommige studenten verwart of schrikt, komt het hierop neer: om een skeletspiervezel te samentrekken, moet het membraan eerst “opgewonden”zijn—met andere woorden, moet het worden gestimuleerd om een actiepotentiaal te ontsteken. De spiervezel actie potentieel, die veegt langs de sarcolemma als een golf, is “gekoppeld” aan de werkelijke samentrekking door de afgifte van calciumionen (Ca++) van de SR. eenmaal vrijgegeven, De Ca++ interactie met de afschermende eiwitten, waardoor ze opzij te bewegen, zodat de actin-bindende plaatsen beschikbaar zijn voor bevestiging door myosine hoofden. Myosin trekt dan de actin filamenten naar het centrum, verkorting van de spiervezel.
in skeletspieren begint deze sequentie met signalen van de somatische motorische deling van het zenuwstelsel. Met andere woorden, de “excitatie” stap in skeletspieren wordt altijd veroorzaakt door het signaleren van het zenuwstelsel (Figuur 4).
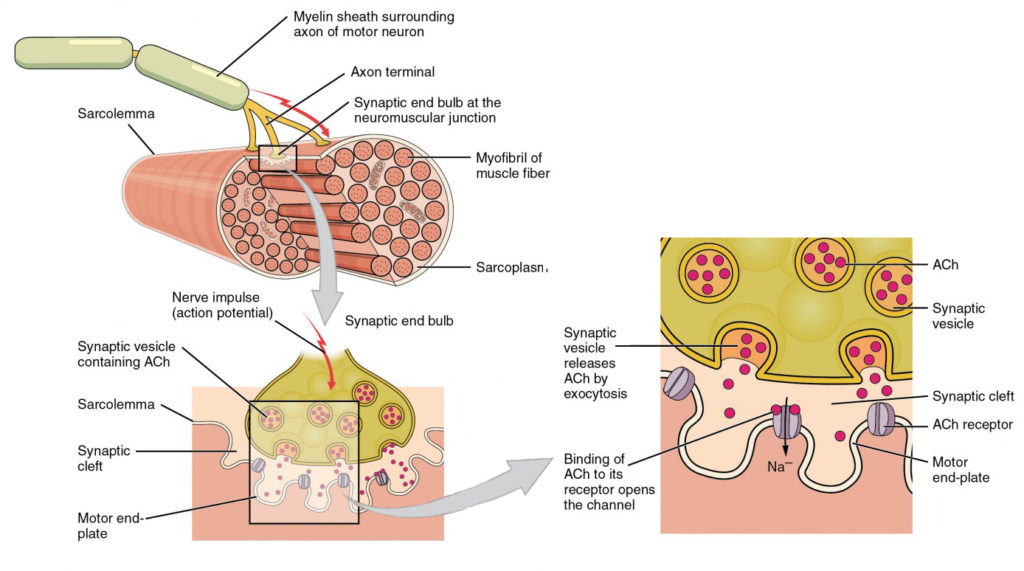
Figuur 4. Motor eindplaat en innervatie. Op de NMJ, de Axon terminal releases ACh. De motoreindplaat is de locatie van de ACh-receptoren in de spiervezel sarcolemma. Wanneer ACh de molecules worden vrijgegeven, diffunderen zij over een minieme ruimte genoemd synaptische gespleten en binden aan de receptoren.
de motorische neuronen die de skeletspiervezels vertellen samen te trekken, ontstaan in het ruggenmerg, met een kleiner aantal in de hersenstam voor activering van skeletspieren van het gezicht, hoofd en nek. Deze neuronen hebben lange processen, genaamd axonen, die gespecialiseerd zijn om actiepotentialen lange afstanden— in dit geval, helemaal van het ruggenmerg naar de spier zelf (die tot drie meter afstand). De axonen van meerdere neuronen bundelen zich samen om zenuwen te vormen, zoals draden die samen in een kabel zijn gebundeld.
Het signaleren begint wanneer een neuronaal actiepotentiaal langs het axon van een motorneuron reist, en dan langs de individuele takken om bij de NMJ te eindigen. Bij de NMJ geeft de Axon terminal een chemische boodschapper, of neurotransmitter, genaamd acetylcholine (ACh). De ACh-moleculen diffunderen over een minieme ruimte, de synaptische gespleten en binden zich aan ACh-receptoren die zich bevinden in de motorische eindplaat van het sarcolemma aan de andere kant van de synaps. Zodra ACh bindt, opent een kanaal in de ACh-receptor en positief geladen ionen kunnen door in de spiervezel gaan, veroorzakend het om te depolariseren, betekenend dat het membraanpotentieel van de spiervezel minder negatief wordt (dichter bij nul.)
naarmate het membraan depolariseert, worden een andere reeks ionenkanalen, genaamd spanningsgesloten natriumkanalen, geactiveerd om te openen. Natriumionen gaan de spiervezel in, en een actiepotentiaal verspreidt zich snel (of “branden”) langs het gehele membraan om excitatie-contractie koppeling in werking te stellen.
dingen gebeuren zeer snel in de wereld van prikkelbare membranen (denk er eens over na hoe snel je met je vingers kunt knippen zodra je besluit om het te doen). Onmiddellijk na depolarisatie van het membraan, repolariseert het, het herstellen van het negatieve membraanpotentieel. Ondertussen wordt ACh in de synaptische spleet afgebroken door het enzym acetylcholinesterase (AChE) zodat ACh niet kan terugkeren naar een receptor en zijn kanaal kan heropenen, wat ongewenste uitgebreide spieropwrijving en samentrekking zou veroorzaken.

Figuur 5. De T-tubule. Smalle t-tubuli maken de geleiding van elektrische impulsen mogelijk. De functies van SR om intracellular niveaus van calcium te regelen. Twee terminale cisternae (waar vergrote SR aansluit op de T-tubule) en een T—tubule bestaan uit een triade-een “trio” van membranen, met die van SR aan twee zijden en de T-tubule ingeklemd tussen hen.
vermeerdering van een actiepotentiaal langs het sarcolemma is het excitatiegedeelte van excitatie-contractiekoppeling. Bedenk dat deze opwinding eigenlijk leidt tot de afgifte van calciumionen (Ca++) uit de opslag in de cel SR. Voor het actiepotentieel om het membraan van de SR te bereiken, zijn er periodieke invaginaties in het sarcolemma, genaamd T-tubuli (“T “staat voor”transversaal”). U zult zich herinneren dat de diameter van een spiervezel tot 100 µm kan zijn, zodat deze T-tubuli ervoor zorgen dat het membraan dicht bij de SR in het sarcoplasma kan komen. De opstelling van een T-tubule met de membranen van SR aan weerszijden wordt een triade genoemd (Figuur 5). De triade omringt de cilindrische structuur genoemd een myofibril, die actin en myosine bevat.
De T-tubuli brengen de actiepotentiaal naar de binnenkant van de cel, waardoor de opening van calciumkanalen in het membraan van de aangrenzende SR wordt geactiveerd, waardoor Ca++ uit de SR naar het sarcoplasma diffundert. Het is de komst van Ca++ in het sarcoplasma dat samentrekking van de spiervezel initieert door zijn contractiele eenheden, of sarcomeren.