Obiettivi di Apprendimento
- Descrivere gli strati di tessuto connettivo imballaggio muscolo scheletrico
- Spiegare come i muscoli lavorano con i tendini per spostare il corpo
- Identificare le aree di fibre muscolari scheletriche
- Descrivere l’accoppiamento eccitazione-contrazione
Il più noto funzionalità del muscolo scheletrico è la sua capacità di contratto e la causa del movimento. I muscoli scheletrici agiscono non solo per produrre movimento, ma anche per fermare il movimento, come resistere alla gravità per mantenere la postura. Sono necessari piccoli aggiustamenti costanti dei muscoli scheletrici per tenere un corpo in posizione verticale o bilanciato in qualsiasi posizione. I muscoli impediscono anche l’eccesso di movimento delle ossa e delle articolazioni, mantenendo la stabilità scheletrica e prevenendo danni o deformazioni alla struttura scheletrica. Le articolazioni possono essere disallineate o dislocate interamente tirando le ossa associate; i muscoli lavorano per mantenere stabili le articolazioni. I muscoli scheletrici si trovano in tutto il corpo alle aperture dei tratti interni per controllare il movimento di varie sostanze. Questi muscoli consentono funzioni, come la deglutizione, la minzione e la defecazione, di essere sotto controllo volontario. I muscoli scheletrici proteggono anche gli organi interni (in particolare gli organi addominali e pelvici) agendo come barriera esterna o scudo per traumi esterni e sostenendo il peso degli organi.
I muscoli scheletrici contribuiscono al mantenimento dell’omeostasi nel corpo generando calore. La contrazione muscolare richiede energia e quando l’ATP viene scomposto, viene prodotto calore. Questo calore è molto evidente durante l’esercizio, quando il movimento muscolare sostenuto fa aumentare la temperatura corporea, e nei casi di freddo estremo, quando tremare produce contrazioni muscolari scheletriche casuali per generare calore.

Figura 1. I tre strati di tessuto connettivo. Fasci di fibre muscolari, chiamati fascicoli, sono coperti dal perimysium. Le fibre muscolari sono coperte dall’endomysium.
Ogni muscolo scheletrico è un organo costituito da vari tessuti integrati. Questi tessuti includono le fibre muscolari scheletriche, i vasi sanguigni, le fibre nervose e il tessuto connettivo. Ogni muscolo scheletrico ha tre strati di tessuto connettivo (chiamato “mysia”) che lo racchiudono e forniscono struttura al muscolo nel suo complesso, e anche compartimentalizzare le fibre muscolari all’interno del muscolo (Figura 1). Ogni muscolo è avvolto in una guaina di tessuto connettivo denso e irregolare chiamato epimysium, che consente a un muscolo di contrarsi e muoversi potentemente mantenendo la sua integrità strutturale. L’epimysium separa anche il muscolo da altri tessuti e organi nell’area, consentendo al muscolo di muoversi indipendentemente.
All’interno di ogni muscolo scheletrico, le fibre muscolari sono organizzate in singoli fasci, ciascuno chiamato un fascicolo, da uno strato intermedio di tessuto connettivo chiamato perimysium. Questa organizzazione fascicolare è comune nei muscoli degli arti; permette al sistema nervoso di innescare un movimento specifico di un muscolo attivando un sottoinsieme di fibre muscolari all’interno di un fascio, o fascicolo del muscolo. All’interno di ogni fascicolo, ogni fibra muscolare è racchiusa in un sottile strato di tessuto connettivo di collagene e fibre reticolari chiamato endomysium. L’endomysium contiene il fluido extracellulare e sostanze nutritive per sostenere la fibra muscolare. Questi nutrienti vengono forniti attraverso il sangue al tessuto muscolare.
Nei muscoli scheletrici che lavorano con i tendini per tirare sulle ossa, il collagene nei tre strati di tessuto (la mysia) si intreccia con il collagene di un tendine. All’altra estremità del tendine, si fonde con il periostio che ricopre l’osso. La tensione creata dalla contrazione delle fibre muscolari viene poi trasferita attraverso la mysia, al tendine, e quindi al periostio per tirare l’osso per il movimento dello scheletro. In altri luoghi, la mysia può fondersi con un ampio foglio tendineo chiamato aponeurosi, o fascia, il tessuto connettivo tra pelle e ossa. L’ampio foglio di tessuto connettivo nella parte bassa della schiena in cui si fondono i muscoli gran dorsale (i “lats”) è un esempio di aponeurosi.
Ogni muscolo scheletrico è anche riccamente fornito dai vasi sanguigni per il nutrimento, la consegna di ossigeno e la rimozione dei rifiuti. Inoltre, ogni fibra muscolare in un muscolo scheletrico è fornita dal ramo assone di un motoneurone somatico, che segnala la fibra a contrarsi. A differenza del muscolo cardiaco e della muscolatura liscia, l’unico modo per contrarre funzionalmente un muscolo scheletrico è attraverso la segnalazione dal sistema nervoso.
Fibre muscolari scheletriche
Poiché le cellule muscolari scheletriche sono lunghe e cilindriche, vengono comunemente chiamate fibre muscolari. Le fibre muscolari scheletriche possono essere abbastanza grandi per le cellule umane, con diametri fino a 100 µm e lunghezze fino a 30 cm (11,8 pollici) nel Sartorio della parte superiore della gamba. Durante lo sviluppo iniziale, i mioblasti embrionali, ciascuno con il proprio nucleo, si fondono con fino a centinaia di altri mioblasti per formare le fibre muscolari scheletriche multinucleate. Nuclei multipli significano copie multiple di geni, permettendo la produzione delle grandi quantità di proteine ed enzimi necessari per la contrazione muscolare.
Qualche altra terminologia associata alle fibre muscolari è radicata nel sarco greco, che significa “carne.”La membrana plasmatica delle fibre muscolari è chiamata sarcolemma, il citoplasma è indicato come sarcoplasma e il reticolo endoplasmatico liscio specializzato, che memorizza, rilascia e recupera ioni calcio (Ca++) è chiamato reticolo sarcoplasmatico (SR) (Figura 2). Come verrà presto descritto, l’unità funzionale di una fibra muscolare scheletrica è il sarcomero, una disposizione altamente organizzata dei miofilamenti contrattili actina (filamento sottile) e miosina (filamento spesso), insieme ad altre proteine di supporto.

Figura 2. Fibra muscolare. Una fibra muscolare scheletrica è circondata da una membrana plasmatica chiamata sarcolemma, che contiene sarcoplasma, il citoplasma delle cellule muscolari. Una fibra muscolare è composta da molte fibrille, che danno alla cellula il suo aspetto striato.
Il Sarcomero
L’aspetto striato delle fibre muscolari scheletriche è dovuto alla disposizione dei miofilamenti di actina e miosina in ordine sequenziale da un’estremità della fibra muscolare all’altra. Ogni pacchetto di questi microfilamenti e le loro proteine regolatrici, troponina e tropomiosina (insieme ad altre proteine) è chiamato sarcomero.
Il sarcomero è l’unità funzionale della fibra muscolare. Il sarcomero stesso è impacchettato all’interno del miofibril che corre l’intera lunghezza della fibra muscolare e si attacca al sarcolemma alla sua estremità. Poiché le miofibrille si contraggono, l’intera cellula muscolare si contrae. Poiché le miofibrille hanno solo un diametro di circa 1,2 µm, si possono trovare da centinaia a migliaia (ciascuna con migliaia di sarcomeri) all’interno di una fibra muscolare. Ogni sarcomero ha una lunghezza di circa 2 µm con una disposizione tridimensionale simile a un cilindro ed è delimitato da strutture chiamate dischi Z (chiamate anche linee Z, perché le immagini sono bidimensionali), a cui sono ancorati i miofilamenti di actina (Figura 3). Poiché l’actina e il suo complesso troponina-tropomiosina (proiettando dai dischi Z verso il centro del sarcomero) formano fili più sottili della miosina, è chiamato il filamento sottile del sarcomero. Allo stesso modo, poiché i fili di miosina e le loro teste multiple (che proiettano dal centro del sarcomero, verso ma non tutti verso, i dischi Z) hanno più massa e sono più spessi, sono chiamati il filamento spesso del sarcomero.

Figura 3. Il sarcomero. Il sarcomero, la regione da una linea Z alla linea Z successiva, è l’unità funzionale di una fibra muscolare scheletrica.
La giunzione neuromuscolare
Un’altra specializzazione del muscolo scheletrico è il sito in cui il terminale di un motoneurone incontra la fibra muscolare—chiamata giunzione neuromuscolare (NMJ). Questo è dove la fibra muscolare prima risponde alla segnalazione da parte del motoneurone. Ogni fibra muscolare scheletrica in ogni muscolo scheletrico è innervata da un motoneurone al NMJ. I segnali di eccitazione dal neurone sono l’unico modo per attivare funzionalmente la fibra per contrarsi.
Accoppiamento eccitazione-contrazione
Tutte le cellule viventi hanno potenziali di membrana, o gradienti elettrici attraverso le loro membrane. L’interno della membrana è di solito intorno -60 a -90 mV, rispetto all’esterno. Questo è indicato come potenziale di membrana di una cellula. I neuroni e le cellule muscolari possono utilizzare i loro potenziali di membrana per generare segnali elettrici. Lo fanno controllando il movimento di particelle cariche, chiamate ioni, attraverso le loro membrane per creare correnti elettriche. Ciò si ottiene aprendo e chiudendo proteine specializzate nella membrana chiamate canali ionici. Sebbene le correnti generate dagli ioni che si muovono attraverso queste proteine del canale siano molto piccole, costituiscono la base sia della segnalazione neurale che della contrazione muscolare.
Sia i neuroni che le cellule muscolari scheletriche sono elettricamente eccitabili, il che significa che sono in grado di generare potenziali d’azione. Un potenziale d’azione è un tipo speciale di segnale elettrico che può viaggiare lungo una membrana cellulare come un’onda. Ciò consente di trasmettere rapidamente e fedelmente un segnale su lunghe distanze.
Anche se il termine accoppiamento eccitazione-contrazione confonde o spaventa alcuni studenti, si riduce a questo: perché una fibra muscolare scheletrica si contragga, la sua membrana deve prima essere “eccitata”—in altre parole, deve essere stimolata a sparare un potenziale d’azione. Il potenziale d’azione della fibra muscolare, che spazia lungo il sarcolemma come un’onda, viene “accoppiato” alla contrazione effettiva attraverso il rilascio di ioni calcio (Ca++) dalla SR. Una volta rilasciato, il Ca++ interagisce con le proteine schermanti, costringendole a spostarsi in modo che i siti di legame con l’actina siano disponibili per il fissaggio da parte delle teste di miosina. La miosina poi tira i filamenti di actina verso il centro, accorciando la fibra muscolare.
Nel muscolo scheletrico, questa sequenza inizia con segnali dalla divisione motoria somatica del sistema nervoso. In altre parole, il passo di “eccitazione” nei muscoli scheletrici viene sempre attivato dalla segnalazione dal sistema nervoso (Figura 4).
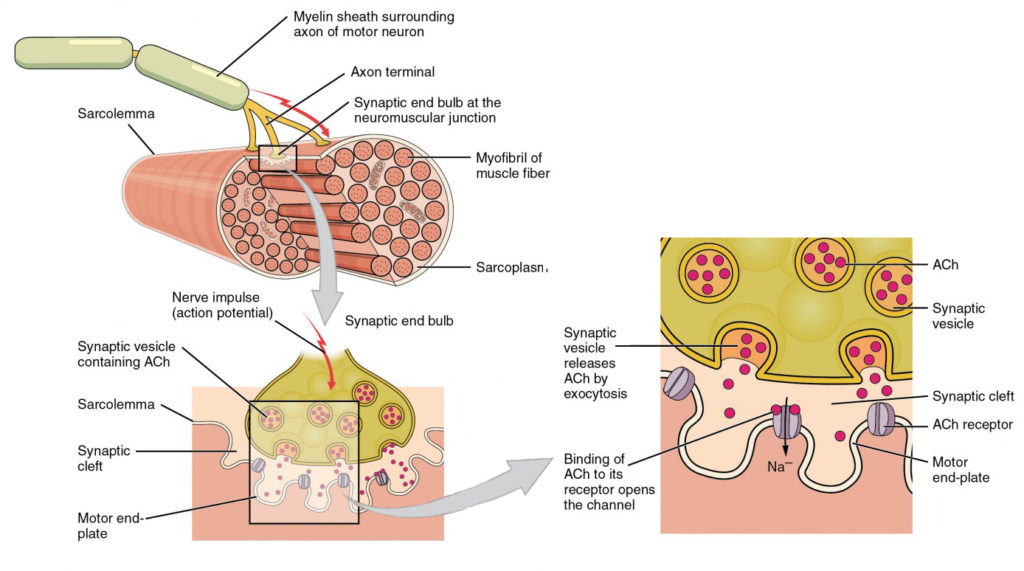
Figura 4. Piastra terminale motore e innervazione. Al NMJ, il terminale axon rilascia ACh. La piastra terminale del motore è la posizione dei recettori ACh nel sarcolemma della fibra muscolare. Quando le molecole ACh vengono rilasciate, si diffondono in uno spazio minuto chiamato fessura sinaptica e si legano ai recettori.
I motoneuroni che dicono alle fibre muscolari scheletriche di contrarsi provengono dal midollo spinale, con un numero minore situato nel tronco cerebrale per l’attivazione dei muscoli scheletrici del viso, della testa e del collo. Questi neuroni hanno processi lunghi, chiamati assoni, che sono specializzati per trasmettere potenziali d’azione lunghe distanze— in questo caso, tutta la strada dal midollo spinale al muscolo stesso (che può essere fino a tre piedi di distanza). Gli assoni di più neuroni si raggruppano per formare nervi, come fili raggruppati in un cavo.
La segnalazione inizia quando un potenziale d’azione neuronale viaggia lungo l’assone di un motoneurone e quindi lungo i singoli rami per terminare al NMJ. Al NMJ, il terminale dell’assone rilascia un messaggero chimico, o neurotrasmettitore, chiamato acetilcolina (ACh). Le molecole di ACh si diffondono in uno spazio minuto chiamato fessura sinaptica e si legano ai recettori ACh situati all’interno della piastra terminale del sarcolemma sull’altro lato della sinapsi. Una volta che ACh si lega, un canale nel recettore ACh si apre e gli ioni caricati positivamente possono passare attraverso la fibra muscolare, facendola depolarizzare, il che significa che il potenziale di membrana della fibra muscolare diventa meno negativo (più vicino a zero.)
Mentre la membrana depolarizza, un altro insieme di canali ionici chiamati canali del sodio voltaggio-gated viene attivato per aprirsi. Gli ioni di sodio entrano nella fibra muscolare e un potenziale d’azione si diffonde rapidamente (o “incendi”) lungo l’intera membrana per avviare l’accoppiamento eccitazione-contrazione.
Le cose accadono molto rapidamente nel mondo delle membrane eccitabili (basti pensare a quanto velocemente si possono schioccare le dita non appena si decide di farlo). Immediatamente dopo la depolarizzazione della membrana, si ripolarizza, ristabilendo il potenziale negativo della membrana. Nel frattempo, l’ACh nella fessura sinaptica viene degradato dall’enzima acetilcolinesterasi (AChE) in modo che l’ACh non possa riconnettersi a un recettore e riaprire il suo canale, il che causerebbe eccitazione e contrazione muscolare estesa indesiderata.

Figura 5. Il tubulo a T. I tubuli a T stretti consentono la conduzione di impulsi elettrici. Le funzioni SR per regolare i livelli intracellulari di calcio. Due cisterne terminali (dove SR allargata collega al T-tubulo) e un T-tubulo comprendono una triade—un “trio” di membrane, con quelli di SR su due lati e il T-tubulo inserita tra di loro.
La propagazione di un potenziale d’azione lungo il sarcolemma è la porzione di eccitazione dell’accoppiamento eccitazione-contrazione. Ricordiamo che questa eccitazione innesca effettivamente il rilascio di ioni calcio (Ca++) dal suo stoccaggio nella SR della cellula. Per il potenziale d’azione per raggiungere la membrana della SR, ci sono invaginazioni periodiche nel sarcolemma, chiamate T-tubuli (“T” sta per “trasversale”). Ricorderai che il diametro di una fibra muscolare può arrivare fino a 100 µm, quindi questi tubuli T assicurano che la membrana possa avvicinarsi alla SR nel sarcoplasma. La disposizione di un tubulo T con le membrane di SR su entrambi i lati è chiamata triade (Figura 5). La triade circonda la struttura cilindrica chiamata miofibril, che contiene actina e miosina.
I tubuli T trasportano il potenziale d’azione all’interno della cellula, che innesca l’apertura dei canali del calcio nella membrana dell’SR adiacente, causando la diffusione di Ca++ dall’SR e nel sarcoplasma. È l’arrivo di Ca++ nel sarcoplasma che avvia la contrazione della fibra muscolare dalle sue unità contrattili, o sarcomeri.