Læringsmål
- Beskriv lagene av bindevevemballasje skjelettmuskulatur
- Forklar hvordan muskler arbeider med sener for å bevege kroppen
- Identifisere områder av skjelettmuskulaturfibrene
- Beskriv eksitasjon-kontraksjonskobling
den mest kjente egenskapen til skjelettmuskulatur er evnen til å trekke seg sammen og forårsake bevegelse. Skjelettmuskler virker ikke bare for å produsere bevegelse, men også for å stoppe bevegelse, for eksempel å motstå tyngdekraften for å opprettholde stillingen. Små, konstante justeringer av skjelettmuskulaturen er nødvendig for å holde en kropp oppreist eller balansert i enhver posisjon. Muskler forhindrer også overflødig bevegelse av bein og ledd, opprettholder skjelettstabilitet og forhindrer skjelettstruktur skade eller deformasjon. Leddene kan bli feiljustert eller dislocated helt ved å trekke på tilhørende bein; muskler arbeider for å holde leddene stabile. Skelettmuskler er plassert i hele kroppen ved åpningene av indre kanaler for å kontrollere bevegelsen av forskjellige stoffer. Disse musklene tillater funksjoner, som svelging, urinering og avføring, å være under frivillig kontroll. Skjelettmuskler beskytter også indre organer (spesielt buk-og bekkenorganer) ved å fungere som en ekstern barriere eller skjold mot ytre traumer og ved å støtte organets vekt.
Skjelettmuskler bidrar til vedlikehold av homeostase i kroppen ved å generere varme. Muskelkontraksjon krever energi, OG NÅR ATP brytes ned, produseres varme. Denne varmen er svært merkbar under trening, når vedvarende muskelbevegelse fører til at kroppstemperaturen stiger, og i tilfeller av ekstrem kulde, når rysting produserer tilfeldige skjelettmuskulaturkontraksjoner for å generere varme.

Figur 1. De Tre Bindevevslagene. Bunter av muskelfibre, kalt fascicles, er dekket av perimysium. Muskelfibre er dekket av endomysium.
hver skjelettmuskulatur er et organ som består av forskjellige integrerte vev. Disse vevene inkluderer skjelettmuskelfibre, blodkar, nervefibre og bindevev. Hver skjelettmuskulatur har tre lag bindevev (kalt «mysia») som omslutter det og gir struktur til muskelen som helhet, og fordeler også muskelfibrene i muskelen (Figur 1). Hver muskel er pakket inn i et skjede av tett, uregelmessig bindevev kalt epimysium, som gjør at en muskel kan trekke seg sammen og bevege seg kraftig samtidig som den opprettholder sin strukturelle integritet. Epimysium skiller også muskler fra andre vev og organer i området, slik at muskelen kan bevege seg selvstendig.innenfor hver skjelettmuskulatur er muskelfibre organisert i individuelle bunter, hver kalt en fascicle, av et mellomlag av bindevev kalt perimysium. Denne fascikulære organisasjonen er vanlig i muskler i lemmer; det gjør at nervesystemet kan utløse en bestemt bevegelse av en muskel ved å aktivere en delmengde av muskelfibre i en bunt eller muskelbjelke. Inne i hver fascicle er hver muskelfiber innkapslet i et tynt bindevevslag av kollagen og retikulære fibre kalt endomysium. Endomysium inneholder ekstracellulær væske og næringsstoffer for å støtte muskelfiberen. Disse næringsstoffene leveres via blod til muskelvevet.
i skjelettmuskler som arbeider med sener å trekke på bein, kollagen i de tre vev lag (mysia) fletter med kollagen av en sene. I den andre enden av senen smelter den sammen med periosteumet som dekker beinet. Spenningen som oppstår ved sammentrekning av muskelfibrene overføres deretter til mysia, til senen, og deretter til periosteum for å trekke på beinet for bevegelse av skjelettet. På andre steder kan mysia smelte sammen med et bredt, senelignende ark kalt en aponeurose, eller til fascia, bindevevet mellom hud og bein. Det brede arket av bindevev i nedre rygg som latissimus dorsi muskler («lats») smelter inn i er et eksempel på en aponeurose.
hver skjelettmuskulatur leveres også rikt av blodkar for næring, oksygentilførsel og fjerning av avfall. I tillegg leveres hver muskelfiber i en skjelettmuskulatur av axongrenen til en somatisk motorneuron, som signalerer fiberen til kontrakt. I motsetning til hjerte og glatt muskulatur, er den eneste måten å funksjonelt kontrakt en skjelettmuskulatur gjennom signalering fra nervesystemet.
Skjelettmuskelfibre
fordi skjelettmuskelceller er lange og sylindriske, blir de ofte referert til som muskelfibre. Skjelettmuskelfibre kan være ganske store for humane celler, med diametre opp til 100 µ og lengder opp til 30 cm (11,8 tommer) I Sartorius i overbenet. Under tidlig utvikling smelter embryonale myoblaster, hver med sin egen kjerne, sammen med opptil hundrevis av andre myoblaster for å danne multinukleerte skjelettmuskelfibre. Flere kjerner betyr flere kopier av gener, som tillater produksjon av store mengder proteiner og enzymer som trengs for muskelkontraksjon.
Noen annen terminologi forbundet med muskelfibre er forankret i den greske sarkoen, som betyr » kjøtt.»Plasmamembranen i muskelfibre kalles sarcolemma, cytoplasma refereres til som sarcoplasma, og det spesialiserte glatte endoplasmatiske retikulumet, som lagrer, frigjør og henter kalsiumioner (Ca++) kalles sarcoplasmatisk retikulum (Sr) (Figur 2). Som det snart vil bli beskrevet, er den funksjonelle enheten av en skjelettmuskelfiber sarkomeren, et høyt organisert arrangement av kontraktile myofilamenter actin (tynn filament) og myosin (tykt filament), sammen med andre støtteproteiner.

figur 2. Muskel Fiber. En skjelettmuskelfiber er omgitt av en plasmamembran kalt sarcolemma, som inneholder sarcoplasma, cytoplasma av muskelceller. En muskel fiber består av mange fibriller, som gir cellen sin tverrstripet utseende.
Sarkomeren
det strierte utseendet av skjelettmuskelfibre skyldes arrangementet av myofilamentene av aktin og myosin i sekvensiell rekkefølge fra den ene enden av muskelfibre til den andre. Hver pakke av disse mikrofilamenter og deres regulatoriske proteiner, troponin og tropomyosin (sammen med andre proteiner) kalles en sarkomere.
sarkomeren er den funksjonelle enheten til muskelfiberen. Sarcomeren selv er buntet i myofibrilen som løper hele lengden av muskelfiberen og festes til sarcolemma ved enden. Som myofibriller kontrakt, kontrakter hele muskelcellen. Fordi myofibriller bare er omtrent 1,2 µ i diameter, kan hundrevis til tusenvis (hver med tusenvis av sarkomerer) finnes i en muskelfiber. Hver sarcomere er omtrent 2 µ i lengde med et tredimensjonalt sylinderlignende arrangement og grenser til strukturer kalt Z-skiver (også kalt Z-linjer, fordi bilder er todimensjonale), som aktinmyofilamentene er forankret til (Figur 3). Fordi aktin og dets troponin-tropomyosin-kompleks (projiserer Fra Z-skivene mot midten av sarkomeren) danner tråder som er tynnere enn myosinet, kalles det sarcomerens tynne filament. På samme måte, fordi myosinstrengene og deres flere hoder (projiserer fra midten av sarkomeren, mot, men ikke alle til vei Til, Z-platene) har mer masse Og er tykkere, kalles de sarkomerens tykke filament.

figur 3. Sarkomeren. Sarcomeren, regionen Fra En Z-linje Til neste Z-linje, er den funksjonelle enheten til en skjelettmuskulaturfiber.
Det Nevromuskulære Krysset
En annen spesialisering av skjelettmuskulaturen er stedet der en motorneurons terminal møter muskelfiberen-kalt DET nevromuskulære krysset (NMJ). Det er her muskelfiberen først reagerer på signalering av motorneuronen. Hver skjelettmuskel fiber i hver skjelettmuskel er innervert av en motor neuron VED NMJ. Excitasjonssignaler fra nevronen er den eneste måten å funksjonelt aktivere fiberen til kontrakt.
Eksitasjon-Sammentrekning Kopling
alle levende celler har membranpotensialer, eller elektriske gradienter over sine membraner. Innsiden av membranen er vanligvis rundt -60 til -90 mV, i forhold til utsiden. Dette kalles cellens membranpotensial. Nevroner og muskelceller kan bruke membranpotensialene til å generere elektriske signaler. De gjør dette ved å kontrollere bevegelsen av ladede partikler, kalt ioner, over deres membraner for å skape elektriske strømmer. Dette oppnås ved å åpne og lukke spesialiserte proteiner i membranen kalt ionkanaler. Selv om strømmene som genereres av ioner som beveger seg gjennom disse kanalproteinene, er svært små, danner de grunnlaget for både nevrale signalering og muskelkontraksjon.både nevroner og skjelettmuskelceller er elektrisk spennende, noe som betyr at de er i stand til å generere handlingspotensialer. Et handlingspotensial er en spesiell type elektrisk signal som kan bevege seg langs en cellemembran som en bølge. Dette gjør at et signal kan overføres raskt og trofast over lange avstander.Selv om begrepet eksitasjonskobling forvirrer eller skremmer noen studenter, kommer det ned til dette: for at en skjelettmuskelfiber skal kontrakt, må membranen først være «begeistret»—med andre ord, det må stimuleres til å brenne et handlingspotensial. Muskelfiberaksjonspotensialet, som feier langs sarcolemma som en bølge, er «koblet» til den faktiske sammentrekningen gjennom frigjøring av kalsiumioner (Ca++) fra SR. Når Det er utgitt, interagerer Ca++ med skjermproteinene, og tvinger dem til å bevege seg til side slik at aktinbindingsstedene er tilgjengelige for vedlegg av myosinhoder. Myosinet trekker deretter aktinfilamentene mot midten, forkorter muskelfiberen.
i skjelettmuskulatur begynner denne sekvensen med signaler fra den somatiske motordelingen i nervesystemet. Med andre ord utløses «excitasjonstrinnet» i skjelettmuskler alltid ved signalering fra nervesystemet(Figur 4).
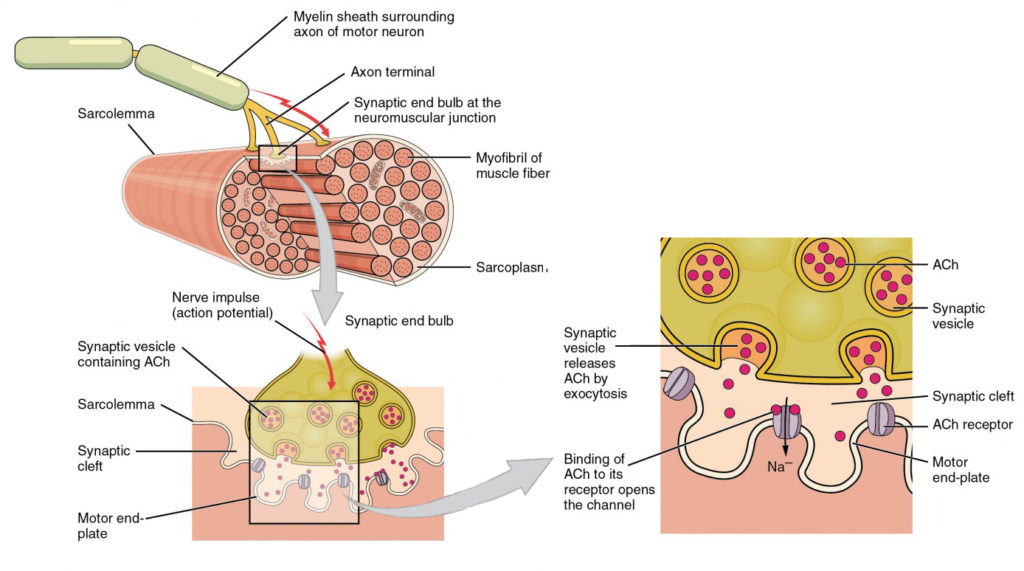
Figur 4. Motor Endeplate og Innervering. På NMJ frigjør axon-terminalen ACh. Motorens endeplate er plasseringen Av ACh-reseptorene i muskelfiberen sarcolemma. Når ach-molekyler frigjøres, diffunderer de over et minuttrom kalt synaptisk kløft og binder seg til reseptorene.motorneuronene som forteller skjelettmuskelfibrene til kontrakt, kommer fra ryggmargen, med et mindre antall plassert i hjernestammen for aktivering av skjelettmuskler i ansikt, hode og nakke. Disse nevronene har lange prosesser, kalt axoner, som er spesialisert på å overføre handlingspotensialer lange avstander— i dette tilfellet helt fra ryggmargen til selve muskelen (som kan være opptil tre meter unna). Axons av flere nevroner bunt sammen for å danne nerver, som ledninger samlet sammen i en kabel.Signalering begynner når et neuronal handlingspotensial beveger seg langs axonen til et motorneuron, og deretter langs de enkelte grenene for å avslutte VED NMJ. VED NMJ frigir axonterminalen en kjemisk budbringer, eller nevrotransmitter, kalt acetylkolin (ach). Ach-molekylene diffunderer over et minuttrom kalt synaptisk kløft og binder Seg til ACh-reseptorer som ligger i motorens endeplate på sarcolemma på den andre siden av synapsen. Når ACh bindes, åpnes en kanal i ACh-reseptoren og positivt ladede ioner kan passere gjennom i muskelfiberen, noe som får den til å depolarisere, noe som betyr at membranpotensialet i muskelfiberen blir mindre negativt (nærmere null.)
når membranen depolariserer, utløses et annet sett med ionkanaler kalt spenningsstyrte natriumkanaler for å åpne. Natriumioner går inn i muskelfiberen, og et handlingspotensial sprer seg raskt (eller «branner») langs hele membranen for å initiere eksitasjons-sammentrekningskobling.
Ting skjer veldig raskt i verden av spennende membraner (bare tenk på hvor raskt du kan knipse fingrene så snart du bestemmer deg for å gjøre det). Umiddelbart etter depolarisering av membranen repolariserer den, gjenoppretter det negative membranpotensialet. I mellomtiden blir ACh i det synaptiske spaltet degradert av enzymet acetylkolinesterase (AChE) slik at ACh ikke kan binde seg til en reseptor og gjenåpne kanalen, noe som vil forårsake uønsket utvidet muskeleksitasjon og sammentrekning.

figur 5. T-tubule. Smale t-tubuli tillater ledning av elektriske impulser. SR fungerer for å regulere intracellulære nivåer av kalsium. To terminal cisternae (hvor forstørret SR kobles Til T-tubule) og en T-tubule omfatter en triade – en «trekant» av membraner, med de AV SR på to sider og T-tubule klemt mellom dem.
Forplantning av et handlingspotensial langs sarcolemma er eksitasjonspartiet av eksitasjons-kontraksjonskobling. Husk at denne eksitasjonen faktisk utløser frigjøring av kalsiumioner (Ca++) fra lagringen i cellens SR. For handlingspotensialet for å nå membranen TIL SR, er det periodiske invaginasjoner i sarcolemma, kalt T-tubuli («T «står for»tverrgående»). Du vil huske at diameteren på en muskelfiber kan være opp til 100 µ, slik at Disse t-tubulene sikrer at membranen kan komme nær SR i sarkoplasma. Arrangementet Av En T-tubule med membranene TIL SR på hver side kalles en triade(Figur 5). Triaden omgir den sylindriske strukturen kalt en myofibril, som inneholder aktin og myosin.
T-tubulene bærer handlingspotensialet inn i det indre av cellen, som utløser åpningen av kalsiumkanaler i membranen til den tilstøtende SR, noe Som forårsaker Ca++ å diffundere UT AV SR og inn i sarkoplasma. Det er ankomsten Av Ca++ i sarkoplasma som initierer sammentrekning av muskelfiberen ved sine kontraktile enheter eller sarkomerer.