jednostki ruchowe
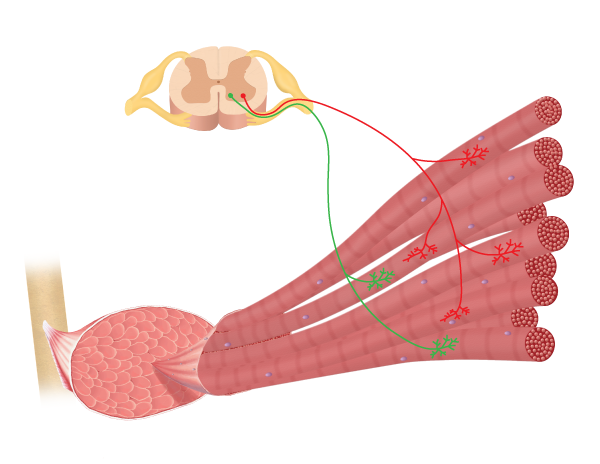
Obraz narysowany przez ucznia BYU-i Nate Shoemaker Wiosna 2016
neurony ruchowe, które unerwiają włókna mięśni szkieletowych, nazywane są neuronami ruchowymi Alfa. Gdy neuron ruchowy Alfa wchodzi w mięsień, dzieli się na kilka gałęzi, z których każda unerwia włókno mięśniowe (zauważ to na powyższym obrazku). Jeden neuron ruchowy Alfa wraz ze wszystkimi włóknami mięśniowymi, które unerwia, jest jednostką motoryczną . Wielkość jednostki motorycznej koreluje z funkcją mięśnia. W mięśniach zaangażowanych w drobną, skoordynowaną kontrolę, jednostki ruchowe są bardzo małe, z 3-5 włóknami mięśniowymi na neuron ruchowy. Mięśnie, które kontrolują ruch oka i mięśnie w naszych rękach mają stosunkowo małe jednostki motoryczne. Z drugiej strony w mięśniach zaangażowanych w silniejsze, ale mniej skoordynowane działania, takie jak mięśnie nóg i pleców, jednostki motoryczne są duże z 1000s włókien mięśniowych na neuron ruchowy.
skurcz mięśni
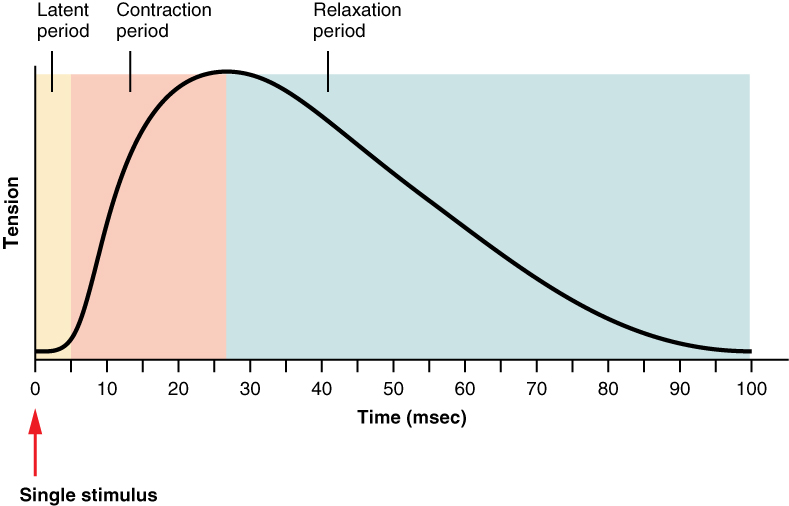
gdy potencjał czynnościowy przemieszcza się w dół neuronu ruchowego, spowoduje to skurcz wszystkich włókien mięśniowych związanych z tym neuronem ruchowym. Skurcz generowany przez pojedynczy potencjał czynnościowy nazywany jest skurczem mięśniowym. Pojedynczy skurcz mięśni składa się z trzech składników. Okres utajony lub Faza opóźnienia, Faza skurczu i faza relaksacji. Okres utajony to krótkie opóźnienie (1-2 milisekundy) od momentu, w którym potencjał czynnościowy dociera do mięśnia, aż do zaobserwowania napięcia w mięśniu. Jest to czas potrzebny do rozproszenia wapnia z SR, wiązania się z troponiną, przemieszczania tropomiozyny z miejsc aktywnych, tworzenia mostków krzyżowych i odbierania wszelkich luzów, które mogą znajdować się w mięśniach. Faza skurczu jest wtedy, gdy mięsień generuje napięcie i jest związany z cyklem mostków krzyżowych, a faza relaksacji to czas powrotu mięśnia do normalnej długości. Długość skurczu różni się w zależności od rodzaju mięśni i może wynosić nawet 10 ms (milisekundy) lub nawet 100 ms (więcej o tym później).
Jeśli drganie mięśni jest tylko jednym szybkim skurczem, po którym następuje natychmiastowe rozluźnienie, jak wyjaśnić płynny ciągły ruch naszych mięśni, gdy kurczą się i poruszają kośćmi w dużym zakresie ruchu? Odpowiedź leży w kolejności odpalania jednostek silnikowych. Jeśli wszystkie jednostki motoryczne wystrzeliły jednocześnie, cały mięsień szybko się kurczy i rozluźnia, powodując bardzo gwałtowny ruch. Zamiast tego, gdy mięsień się kurczy, jednostki motoryczne strzelają asynchronicznie, to znaczy, jeden się kurczy, a następnie ułamek sekundy później inny się kurczy, zanim pierwszy będzie miał czas na relaks, a następnie inny odpali i tak dalej. Tak więc zamiast szybkiego, szarpanego ruchu cały skurcz mięśni jest bardzo płynny i kontrolowany. Nawet gdy mięsień jest w spoczynku, jest losowe odpalanie jednostek motorycznych. To przypadkowe wypalanie jest odpowiedzialne za to, co jest znane jako napięcie mięśniowe. Tak więc mięsień nigdy nie jest” całkowicie ” zrelaksowany, nawet podczas snu. Jeśli jednak neuron do mięśnia zostanie przecięty, nie będzie „napięcia mięśniowego”, co nazywa się porażeniem wiotkim. Istnieje kilka zalet napięcia mięśniowego: najpierw zajmuje ” luz ” w mięśniu, aby gdy zostanie poproszony o kurczenie się, mógł natychmiast zacząć generować napięcie i poruszać kończyną. Jeśli kiedykolwiek holowałeś samochód, wiesz, co się stanie, jeśli nie wyjmiesz luzu z liny holowniczej przed rozpoczęciem ciągnięcia. Drugą rzeczą, którą robi napięcie mięśniowe, jest powstrzymanie zaniku mięśni.
rodzaje skurczów mięśni
skurcze mięśni są opisane na podstawie dwóch zmiennych: Siła (napięcie) i długość (skrócenie). Gdy napięcie w mięśniu wzrasta bez odpowiedniej zmiany długości, skurcz nazywa się skurczem izometrycznym (iso = taki sam, metryczny = długość). Skurcze izometryczne są ważne w utrzymaniu postawy lub stabilizacji stawu. Z drugiej strony, jeśli długość mięśni zmienia się, podczas gdy napięcie mięśni pozostaje względnie stałe, to skurcz nazywa się skurczem izotonicznym (tonik = napięcie). Ponadto skurcze izotoniczne można klasyfikować na podstawie zmian długości. Jeśli mięsień generuje napięcie i cały mięsień się skraca, to jest to skurcz koncentryczny. Przykładem może być zwijanie ciężaru z talii do ramienia; mięsień biceps użyty do tego ruchu uległby koncentrycznemu skurczowi. W przeciwieństwie do tego, przy obniżaniu ciężaru z ramienia do talii biceps również generowałby siłę, ale mięsień wydłużyłby się, jest to skurcz mimośrodowy. Skurcze mimośrodowe działają w celu spowolnienia ruchu w stawie. Dodatkowo, skurcze mimośrodowe mogą generować większą siłę niż skurcze koncentryczne. Pomyśl o dużym pudełku, które zdejmujesz z górnej półki swojej szafy. Można go obniżyć pod całkowitą kontrolą za pomocą mimośrodowych skurczów, ale kiedy próbujesz go zwrócić na półkę za pomocą koncentrycznych skurczów, nie możesz wytworzyć wystarczającej siły, aby podnieść go z powrotem. Trening siłowy, obejmujące zarówno koncentrycznych i ekscentrycznych skurczów, wydaje się zwiększyć siłę mięśni więcej niż tylko koncentrycznych skurczów sam. Jednak ekscentryczne skurcze powodują więcej uszkodzeń (łzawienie) mięśni, co powoduje większy ból mięśni. Jeśli kiedykolwiek biegać w dół w długim wyścigu, a następnie doświadczył bolesność mięśni czworogłowych następnego dnia, wiesz, o czym mówimy.
wielkość mięśni zależy od liczby i wielkości miofibryli, co z kolei zależy od ilości białek włóknistych. Tak więc, trening oporowy indukuje kaskadę zdarzeń, które prowadzą do produkcji większej ilości białek. Często jest to inicjowane przez małe, mikro-łzy w i wokół włókien mięśniowych. Jeśli łzawienie wystąpi na poziomie miofibrilu, mięsień zareaguje zwiększając ilość białek, wzmacniając i powiększając mięsień, zjawisko zwane przerostem. Uważa się, że to łzawienie odpowiada za bolesność mięśni, której doświadczamy po treningu. Jak wspomniano powyżej, naprawa tych małych łez powoduje powiększenie włókien mięśniowych, ale także powoduje wzrost ilości tkanki łącznej w mięśniu. Kiedy osoba „pęka” z treningu siłowego, znaczny procent wzrostu wielkości mięśni jest spowodowany wzrostem ilości tkanki łącznej. Należy podkreślić, że trening wytrzymałościowy nie powoduje znacznego wzrostu wielkości mięśni, ale zwiększa jego zdolność do wytwarzania ATP aerobicznie.
czynniki wpływające na siłę skurczu mięśni
oczywiście nasze mięśnie są w stanie generować różne poziomy siły podczas całego skurczu mięśni. Niektóre działania wymagają znacznie więcej siły generowania niż inne; pomyśl o podniesieniu ołówka w porównaniu z podniesieniem wiadra wody. Powstaje pytanie, jak można generować różne poziomy siły?
sumowanie jednostek motorycznych lub Rekrutacja: wspomniano wcześniej, że wszystkie jednostki motoryczne w mięśniu zwykle nie strzelają w tym samym czasie. Jednym ze sposobów na zwiększenie ilości generowanej siły jest zwiększenie liczby jednostek silnikowych, które strzelają w danym czasie. Mówimy, że rekrutuje się więcej jednostek motorycznych. Im większe obciążenie staramy się przenieść tym więcej jednostek silnikowych, które są aktywowane. Jednak nawet podczas generowania maksymalnej możliwej siły, jesteśmy w stanie użyć tylko około 1/3 naszych całkowitych jednostek silnikowych w jednym czasie. Zwykle będą strzelać asynchronicznie, aby wygenerować maksymalną siłę i zapobiec zmęczeniu mięśni. Gdy włókna zaczynają się męczyć, są zastępowane przez inne w celu utrzymania siły. Bywają jednak chwile, kiedy w ekstremalnych warunkach jesteśmy w stanie zwerbować jeszcze więcej jednostek motorowych. Słyszałeś historie matek podnoszących samochody swoich dzieci, może to nie być całkowicie fikcja. Obejrzyj poniższy klip, aby zobaczyć, jak niesamowite może być ludzkie ciało. Rekrutacja mięśni. (Transkrypcja wideo dostępna)
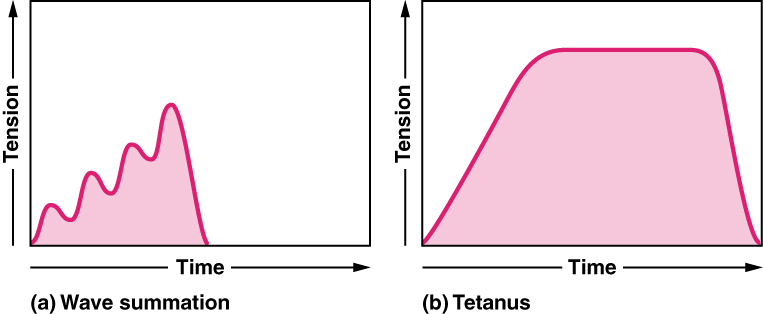
sumowanie fal: Przypomnijmy, że drgania mięśni mogą trwać do 100 ms, a potencjał działania trwa tylko 1-2 ms. ponadto, w przypadku drgań mięśni nie ma okresu oporności, więc można je ponownie stymulować w dowolnym momencie. Jeśli miałbyś stymulować pojedynczą jednostkę motoryczną o coraz wyższych częstotliwościach potencjałów działania, zauważyłbyś stopniowy wzrost siły generowanej przez ten mięsień. Zjawisko to nazywa się sumowaniem fal. Ostatecznie częstotliwość potencjałów działania byłaby tak wysoka, że nie byłoby czasu na rozluźnienie mięśni między kolejnymi bodźcami i pozostałby całkowicie zakontraktowany, stan zwany tężcem. Zasadniczo przy wysokiej częstotliwości działania potencjałów nie ma czasu na usunięcie wapnia z cytozolu. Następnie siła maksymalna jest generowana z maksymalną rekrutacją i częstotliwością potencjału czynnościowego wystarczającą do wywołania tężca.
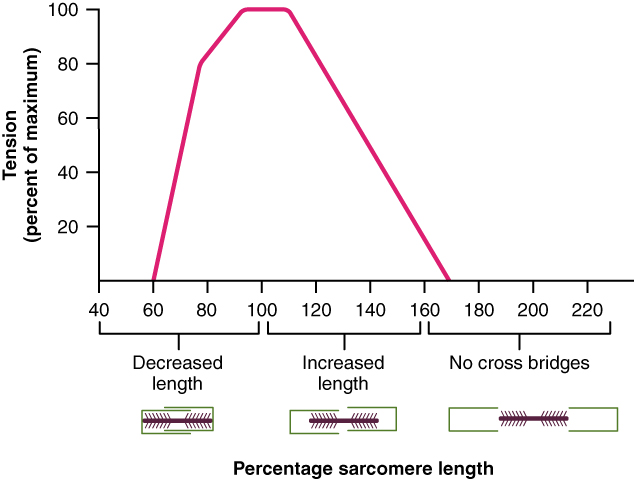
początkowa długość Sarkomeru: Wykazano eksperymentalnie, że początkowa długość sarkomeru wpływa na ilość siły, jaką może wygenerować mięsień. Ta obserwacja ma związek z nakładaniem się grubych i cienkich włókien. Jeśli początkowa długość sarkomeru jest bardzo krótka, grube włókna będą już naciskać na dysk Z i nie ma możliwości dalszego skracania sarkomeru, a mięsień nie będzie w stanie wygenerować takiej siły. Z drugiej strony, jeśli mięsień jest rozciągnięty do punktu, w którym głowy miozyny nie mogą już kontaktować się z aktyną, to znowu wygeneruje się mniej siły. Maksymalna siła jest generowana, gdy mięsień jest rozciągnięty do punktu, który pozwala każdej głowie miozyny na kontakt z aktyną, a sarcomere ma maksymalną odległość do skrócenia. Innymi słowy, grube włókna znajdują się na samych końcach cienkich włókien. Dane te zostały wygenerowane eksperymentalnie przy użyciu mięśni żaby, które zostały rozcięte i rozciągnięte między dwoma prętami. Nienaruszone mięśnie w naszym ciele nie są zwykle rozciągnięte zbyt daleko poza ich optymalną długość ze względu na rozmieszczenie załączników mięśniowych i stawów.
Możesz jednak zrobić mały eksperyment, który pomoże Ci zobaczyć, jak traci się siłę, gdy mięsień jest w bardzo krótkiej lub bardzo rozciągniętej pozycji. W tym eksperymencie wykorzystamy mięśnie, które pomogą Ci przypiąć pad kciuka do klocków palców. Mięśnie te są w pobliżu maksymalnego rozciągnięcia, gdy wyciągniesz rękę, a także wydłużysz nadgarstek. Jak Twój nadgarstek jest cocked powrotem do maksymalnego przedłużenia, spróbuj uszczypnąć kciuk do palców. Widzisz jakie to uczucie słabe? Teraz stopniowo wyginaj nadgarstek do pozycji prostej lub neutralnej. Powinieneś poczuć, jak twoje szczypanie staje się silniejsze. Zgnij łokieć i nadgarstek. Z nadgarstkiem w maksymalnym zgięciu, mięśnie, których używasz do szczypania, są w pobliżu ich najbardziej skróconej pozycji. Spróbuj jeszcze raz szczypać. Powinno być słabe. Ale, ponownie, jak przedłużyć nadgarstek z powrotem do neutralnego należy czuć szczypta się silniejszy.
źródłem energii dla skurczu mięśni
ostatecznym źródłem energii dla skurczu mięśni jest ATP. Przypomnijmy, że każdy cykl głowy miozyny wymaga cząsteczki ATP. Pomnóż to przez wszystkie głowy miozyny w mięśniu i liczbę cykli, które każda głowa kończy każde drganie, a możesz zacząć widzieć, ile ATP jest potrzebne do funkcjonowania mięśni. Szacuje się, że spalamy około całej masy ciała w ATP każdego dnia, więc staje się oczywiste, że musimy stale uzupełniać to ważne źródło energii. W przypadku skurczu mięśni istnieją cztery sposoby, w jakie nasze mięśnie otrzymują ATP wymagane do skurczu.
- CYTOSOLIC ATP: ten ATP reprezentuje „pływającą” pulę ATP lub tę, która jest obecna i dostępna w cytoplazmie. Ten ATP nie wymaga tlenu (beztlenowego), aby go (ponieważ już tam jest) i jest natychmiast dostępny, ale jest krótkotrwały. Zapewnia wystarczającą ilość energii na kilka sekund maksymalnej aktywności w mięśniach-nie jest najlepszym źródłem długotrwałego skurczu. Niemniej jednak, dla mięśni oczu, które stale kurczą się szybko, ale przez krótki czas, jest to świetne źródło.
- fosforan kreatyny: gdy zapasy CYTOZOLICZNE ATP są wyczerpane, komórka wzywa inne szybkie źródło energii, fosforan kreatyny. Fosforan kreatyny jest wysokoenergetycznym związkiem, który może szybko przenieść swój fosforan do cząsteczki ADP, aby szybko uzupełnić ATP bez użycia tlenu. Ten transfer wymaga enzymu kinazy kreatynowej, enzymu, który znajduje się na linii m sarkomere. Fosforan kreatyny może kilkakrotnie uzupełniać pulę ATP, wystarczająco, aby przedłużyć skurcz mięśni do około 10 sekund. Fosforan kreatyny jest najczęściej stosowanym suplementem przez ciężarowców. Chociaż wykazano pewne korzyści, większość z nich jest bardzo niewielka i ogranicza się do wysoce selektywnych działań.
- glikoliza: Glikoliza, jak sama nazwa wskazuje, jest rozpadem glukozy. Podstawowym źródłem glukozy w tym procesie jest glikogen, który jest przechowywany w mięśniach. Glikoliza może funkcjonować w przypadku braku tlenu i jako taka jest głównym źródłem produkcji ATP podczas aktywności beztlenowej. Ta seria reakcji chemicznych będzie głównym celem w następnej jednostce. Chociaż glikoliza jest bardzo szybka i może dostarczać energii do intensywnej aktywności mięśni, może być utrzymywana tylko przez około minutę, zanim mięśnie zaczną się męczyć.
- oddychanie tlenowe lub oksydacyjne: Mechanizmy wymienione powyżej może dostarczyć ATP może nieco ponad minutę, zanim zmęczenie ustawia się. Oczywiście angażujemy się w aktywność mięśni, która trwa znacznie dłużej niż minutę (spacery, jogging lub jazda na rowerze). Działania te wymagają stałej podaży ATP. Gdy wymagane są ciągłe dostawy ATP, komórki wykorzystują mechanizmy metaboliczne mieszczące się w mitochondriach, które wykorzystują tlen. Zwykle nazywamy te procesy metabolizmem tlenowym lub metabolizmem oksydacyjnym. Wykorzystując te procesy tlenowe, mitochondria mogą dostarczyć wystarczającą ilość ATP do zasilania komórek mięśniowych przez wiele godzin. Wadą metabolizmu tlenowego jest to, że jest wolniejszy niż mechanizmy beztlenowe i nie jest wystarczająco szybki do intensywnej aktywności. Jednak w przypadku umiarkowanego poziomu aktywności działa świetnie. Chociaż glukoza może być również wykorzystywana w metabolizmie tlenowym, wybranym składnikiem odżywczym są kwasy tłuszczowe. Jak opisano poniżej, wolno-drgające i szybko-drgające włókna oksydacyjne są zdolne do wykorzystania metabolizmu tlenowego
zmęczenie
Kiedy myślimy o zmęczeniu mięśni szkieletowych, często używamy słowa zmęczenie, jednak fizjologiczne przyczyny zmęczenia znacznie się różnią. Na najprostszym poziomie zmęczenie jest używane do opisania stanu, w którym mięsień nie jest już w stanie optymalnie się kurczyć. Aby ułatwić dyskusję, podzielimy zmęczenie na dwie szerokie kategorie: zmęczenie centralne i zmęczenie obwodowe. Centralne zmęczenie opisuje niewygodne uczucia, które pochodzą z bycia zmęczonym, jest często nazywany ” zmęczenie psychiczne.”Zasugerowano, że centralne zmęczenie wynika z czynników uwalnianych przez mięsień podczas ćwiczeń, które sygnalizują mózgowi” odczuwanie ” zmęczenia. Zmęczenie psychiczne poprzedza zmęczenie obwodowe i występuje na długo zanim włókno mięśniowe nie może się już kurczyć. Jednym z efektów szkolenia jest nauczenie się, jak przezwyciężyć zmęczenie psychiczne. Podczas treningu dowiadujemy się, że te uczucia nie są tak złe i że możemy kontynuować wykonywanie nawet wtedy, gdy czujesz się niekomfortowo. Z tego powodu elitarni sportowcy zatrudniają trenerów, którzy pchają ich i zmuszają do przejścia obok psychicznego zmęczenia.
obwodowe zmęczenie może wystąpić w dowolnym miejscu między węzłem nerwowo-mięśniowym a elementami kurczliwymi mięśnia. Można go podzielić na dwie podkategorie, zmęczenie o niskiej częstotliwości (bieg maratoński) i wysokiej częstotliwości (trening obwodowy). Zmęczenie wysokiej częstotliwości wynika z upośledzonej pobudliwości błony w wyniku nierównowagi jonów. Potencjalnymi przyczynami są nieodpowiednie funkcjonowanie pompy Na+/K+, późniejsza inaktywacja Kanałów Na+ i upośledzenie kanałów Ca2+. Mięśnie mogą szybko odzyskać siły, zwykle w ciągu 30 minut lub mniej, po zmęczeniu o wysokiej częstotliwości. Zmęczenie o niskiej częstotliwości jest skorelowane z upośledzonym uwalnianiem Ca2+, prawdopodobnie z powodu problemów ze skurczem sprzężenia wzbudzenia. Znacznie trudniej jest odzyskać zmęczenie o niskiej częstotliwości, biorąc od 24 godzin do 72 godzin.
ponadto istnieje wiele innych potencjalnych czynników powodujących zmęczenie, należą do nich: akumulacja fosforanów nieorganicznych, akumulacja jonów wodorowych i następująca zmiana pH, zubożenie glikogenu i nierównowaga w K+. Należy pamiętać, że czynniki, które nie są na liście są ATP i kwas mlekowy, z których oba nie przyczyniają się do zmęczenia. Rzeczywistość jest taka, że nadal nie wiemy dokładnie, co powoduje zmęczenie, a wiele badań jest obecnie poświęconych temu tematowi.
typy włókien mięśni szkieletowych
klasycznie włókna mięśni szkieletowych można sklasyfikować zgodnie z ich szybkością skurczu i odpornością na zmęczenie. Klasyfikacje te są w trakcie weryfikacji, ale podstawowe typy obejmują:
- oksydacyjne włókna mięśniowe o powolnym skurczu (typ I),
- oksydacyjno-glikolityczne włókna mięśniowe o szybkim skurczu (Typ IIA) i
- szybkolityczne włókna glikolityczne (Typ IIX).
włókna szybko drgające (typ II) rozwijają napięcie dwa do trzech razy szybciej niż włókna wolno drgające (typ I). Jak szybko włókno może się kurczyć, jest związane z tym, jak długo trwa Ukończenie cyklu cross-bridge. Zmienność ta wynika z różnych odmian cząsteczek miozyny i tego, jak szybko mogą hydrolizować ATP. Przypomnijmy, że to głowa miozyny dzieli ATP. Włókna szybko drgające mają szybszą zdolność ATPazy (dzielenia ATP na ADP + Pi). Szybko drgające włókna bardzo szybko pompują jony Ca2+ z powrotem do retikulum sarkoplazmatycznego, więc komórki te mają znacznie szybsze drgania niż wolniejsza odmiana. W ten sposób włókna szybko drgające mogą wykonywać wiele skurczów znacznie szybciej niż włókna wolno drgające. Pełna lista włókien mięśniowych różni się od siebie pod względem odporności na zmęczenie znajduje się w poniższej tabeli:
| Slow Twitch Oxidative (Type I) | Fast-twitch Oxidative (Type IIA) | Fast-Twitch Glycolytic (Type IIX) | |
| Myosin ATPase activity | slow | fast | fast |
| Size (diameter) | small | medium | large |
| Duration of contraction | long | short | short |
| SERCA pump activity | slow | fast | fast |
| Fatigue | resistant | resistant | easily fatigued |
| Energy utilization | aerobic/oxidative | both | anerobic/glycolytic |
| capillary density | high | medium | low |
| mitochondria | high numbers | medium numbers | low numbers |
| Color | red (contain myoglobin) | red (contain myoglobin) | white (no myoglobin) |
In human skeletal muscles, the ratio of the various fiber types differs from mięśnie do mięśni. Na przykład mięsień gastrocnemius łydki zawiera około połowy wolnych i pół szybkich włókien typu, podczas gdy głębszy mięsień łydki, soleus, jest głównie powolnym drganiem. Z drugiej strony mięśnie oka są głównie szybkie drgania. W rezultacie mięsień gastrocnemius jest używany w sprincie, podczas gdy mięsień soleus jest ważny dla pozycji stojącej. Ponadto kobiety wydają się mieć wyższy stosunek powolnego Twitcha do szybkiego Twitcha w porównaniu do mężczyzn. „Preferowanym” typem włókna dla sportowców sprinterskich jest glikolityczny szybko drgający, który jest bardzo szybki, jednak większość ludzi ma bardzo niski procent tych włókien, < 1%. Biopsje mięśni jednego sprintera światowej klasy wykazały 72% włókien szybko drgających i zadziwiająco 20% były typu IIX. Święty Graal badań mięśni jest określenie, jak zmienić włókna mięśni szkieletowych z jednego typu do drugiego. Wydaje się, że typy włókien mięśniowych są określane embriologicznie przez typ neuronu, który unerwia włókno mięśniowe. Domyślne mięśnie wydają się być wolne, włókna typu I. Jeśli mięsień jest unerwiony przez mały neuron, włókno mięśniowe pozostanie powolne, podczas gdy duże zmylone włókna indukują szybkie izoformy. Ponadto częstotliwość wypalania neuronu zmienia również Typ włókien mięśniowych. Badania sugerują, że ludzie mają podtypy włókien, stanowiące około <5% mięśni, które są podwójnie unerwione i pozwalają na przełączanie się między powolnym a szybkim. Ogólnie rzecz biorąc, wydaje się, że genetyka określa rodzaj unerwienia, które występuje, a następnie typy włókien mięśniowych i że trening może być w stanie nieznacznie zmienić proporcje ze względu na podwójnie unerwione mięśnie. Ponieważ jednak<5% ma podwójne unerwienie, genetyka będzie odgrywać znacznie większą rolę w typach włókien niż trening.
**możesz użyć poniższych przycisków, aby przejść do następnego lub poprzedniego czytania w tym Module**
