cele uczenia się
- opisz warstwy tkanki łącznej pakujące mięśnie szkieletowe
- wyjaśnij, jak mięśnie pracują ze ścięgnami, aby poruszać ciałem
- Zidentyfikuj obszary włókien mięśni szkieletowych
- opisz połączenie pobudzenie-skurcz
najbardziej znaną cechą mięśnia szkieletowego jest jego zdolność do kurczenia się i spowodować ruch. Mięśnie szkieletowe działają nie tylko w celu wytworzenia ruchu, ale także w celu zatrzymania ruchu, na przykład w celu oporu grawitacji w celu utrzymania postawy. Małe, stałe regulacje mięśni szkieletowych są potrzebne, aby utrzymać ciało w pozycji pionowej lub zrównoważonej w dowolnej pozycji. Mięśnie zapobiegają również nadmiernemu ruchowi kości i stawów, utrzymując stabilność szkieletu i zapobiegając uszkodzeniom lub deformacji struktury szkieletowej. Stawy mogą stać się niewspółosiowy lub przemieszczenie całkowicie przez ciągnięcie na związane kości; mięśnie pracują, aby utrzymać stawy stabilne. Mięśnie szkieletowe znajdują się w całym ciele na otworach wewnętrznych dróg, aby kontrolować ruch różnych substancji. Mięśnie te umożliwiają funkcje, takie jak połykanie, oddawanie moczu i wypróżnianie, być pod dobrowolną kontrolą. Mięśnie szkieletowe chronią również narządy wewnętrzne (zwłaszcza brzuszne i miedniczne), działając jako zewnętrzna bariera lub osłona przed urazami zewnętrznymi i wspierając ciężar narządów.
mięśnie szkieletowe przyczyniają się do utrzymania homeostazy w organizmie poprzez wytwarzanie ciepła. Skurcz mięśni wymaga energii, a po rozbiciu ATP Wytwarza się ciepło. To ciepło jest bardzo zauważalne podczas ćwiczeń, gdy trwały ruch mięśni powoduje wzrost temperatury ciała, aw przypadkach ekstremalnego zimna, gdy dreszcze powodują przypadkowe skurcze mięśni szkieletowych w celu wytworzenia ciepła.

Rysunek 1. Trzy Warstwy Tkanki Łącznej. Wiązki włókien mięśniowych, zwane faszkami, są pokryte przez perymizjum. Włókna mięśniowe pokryte są endomysium.
każdy mięsień szkieletowy jest organem, który składa się z różnych zintegrowanych tkanek. Tkanki te obejmują włókna mięśni szkieletowych, naczynia krwionośne, włókna nerwowe i tkankę łączną. Każdy mięsień szkieletowy ma trzy warstwy tkanki łącznej (zwane „mysia”), które go otaczają i zapewniają strukturę mięśniowi jako całości, a także oddzielają włókna mięśniowe w mięśniu (ryc. 1). Każdy mięsień jest owinięty w powłokę gęstej, nieregularnej tkanki łącznej zwanej epimysium, która pozwala mięśniowi kurczyć się i poruszać się silnie, zachowując jego integralność strukturalną. Epimysium oddziela również mięsień od innych tkanek i narządów w okolicy, umożliwiając mu samodzielne poruszanie się.
wewnątrz każdego mięśnia szkieletowego włókna mięśniowe są zorganizowane w pojedyncze wiązki, z których każdy nazywany jest fascyną, przez środkową warstwę tkanki łącznej zwaną perimysium. Ta organizacja faszyczna jest powszechna w mięśniach kończyn; pozwala układowi nerwowemu na wyzwalanie określonego ruchu mięśnia poprzez aktywację podzbioru włókien mięśniowych w wiązce lub fascynacji mięśniem. Wewnątrz każdej fascynacji, każde włókno mięśniowe jest zamknięte w cienkiej warstwie tkanki łącznej kolagenu i włókien siateczkowych zwanych endomysium. Endomysium zawiera płyn zewnątrzkomórkowy i składniki odżywcze wspierające włókno mięśniowe. Te składniki odżywcze są dostarczane przez krew do tkanki mięśniowej.
w mięśniach szkieletowych, które pracują ze ścięgnami do ciągnięcia kości, kolagen w trzech warstwach tkanki (mysia) przeplata się z kolagenem ścięgna. Na drugim końcu ścięgna łączy się z okostną pokrywającą kość. Napięcie powstałe w wyniku skurczu włókien mięśniowych jest następnie przenoszone przez mysię, do ścięgna, a następnie do okostnej, aby pociągnąć za kość w celu ruchu szkieletu. W innych miejscach mysia może łączyć się z szeroką, ścięgnistą blachą zwaną rozcięgnem lub powięzią, tkanką łączną między skórą a kośćmi. Szeroki arkusz tkanki łącznej w dolnej części pleców, że najszerszy grzbiet mięśni („lats”) fuse do jest przykładem rozcięgna.
każdy mięsień szkieletowy jest również bogato zaopatrywany w naczynia krwionośne w celu odżywiania, dostarczania tlenu i usuwania odpadów. Ponadto każde włókno mięśniowe w mięśniu szkieletowym jest dostarczane przez gałąź aksonu somatycznego neuronu ruchowego, który sygnalizuje włókno do kurczenia się. W przeciwieństwie do mięśnia sercowego i gładkiego, jedynym sposobem na funkcjonalne kurczenie się mięśnia szkieletowego jest sygnalizacja z układu nerwowego.
włókna mięśni szkieletowych
ponieważ komórki mięśni szkieletowych są długie i cylindryczne, są one powszechnie określane jako włókna mięśniowe. Włókna mięśni szkieletowych mogą być dość duże dla komórek ludzkich, o średnicy do 100 µm i długości do 30 cm (11,8 cala) w Sartorius górnej części nogi. Podczas wczesnego rozwoju zarodkowe mioblasty, każdy z własnym jądrem, łączą się z setkami innych mioblastów, tworząc wielojądrzaste włókna mięśni szkieletowych. Wiele jąder oznacza wiele kopii genów, umożliwiając produkcję dużych ilości białek i enzymów potrzebnych do skurczu mięśni.
niektóre inne terminy związane z włóknami mięśniowymi są zakorzenione w greckim sarco, co oznacza „ciało.”Błona plazmatyczna włókien mięśniowych nazywa się sarcolemma, cytoplazma jest określana jako sarkoplazma, a wyspecjalizowane gładkie retikulum endoplazmatyczne, które przechowuje, uwalnia i pobiera jony wapnia (Ca++), nazywa się retikulum sarkoplazmatyczne (SR) (Rysunek 2). Jak wkrótce zostanie opisane, jednostką funkcjonalną włókna mięśni szkieletowych jest sarkomere, wysoce zorganizowany układ kurczliwych włókien mięśniowych aktyny (cienkie włókno) i miozyny (grube włókno), wraz z innymi białkami nośnymi.

Rysunek 2. Włókno Mięśniowe. Włókno mięśni szkieletowych jest otoczone błoną plazmatyczną zwaną sarcolemma, która zawiera sarkoplazmę, cytoplazmę komórek mięśniowych. Włókno mięśniowe składa się z wielu włókien, które nadają komórce Prążkowany wygląd.
Sarkomere
Prążkowany wygląd włókien mięśni szkieletowych jest spowodowany ułożeniem włókien mięśniowych aktyny i miozyny w kolejności od jednego końca włókna mięśniowego do drugiego. Każdy pakiet tych mikrofilamentów i ich białek regulatorowych, troponiny I tropomiozyny (wraz z innymi białkami) nazywa się sarcomere.
sarcomere to jednostka funkcjonalna włókna mięśniowego. Sam sarkomere jest wiązany w obrębie miofibrilu, który biegnie przez całą długość włókna mięśniowego i przyczepia się do sarkolemmy na jej końcu. Gdy miofibryle się kurczą, Cała komórka mięśniowa się kurczy. Ponieważ miofibryle mają tylko około 1,2 µm średnicy, setki do tysięcy (każdy z tysiącami sarkomerów) można znaleźć wewnątrz jednego włókna mięśniowego. Każdy sarkomere ma około 2 µm Długości z trójwymiarowym cylindrycznym układem i jest ograniczony strukturami zwanymi z-dyskami (zwanymi również z-liniami, ponieważ obrazy są dwuwymiarowe), do których zakotwiczone są włókna mięśniowe aktyny (ryc. 3). Ponieważ aktyna i jej kompleks troponinowo-tropomiozyny (wystające z dysków Z w kierunku środka sarkomeru) tworzą pasma, które są cieńsze niż miozyna, nazywa się je cienkim włóknem sarkomeru. Podobnie, ponieważ nici miozyny i ich wielokrotne głowy (wystające od środka sarkomeru, w kierunku, ale nie do końca, do dysków Z) mają większą masę i są grubsze, nazywane są grubym włóknem sarkomeru.

Rysunek 3. Sarkomere. Sarkomere, region od jednej linii Z do następnej linii Z, jest jednostką funkcjonalną włókna mięśni szkieletowych.
Złącze nerwowo—mięśniowe
inną specjalizacją mięśnia szkieletowego jest miejsce, w którym terminal neuronu ruchowego styka się z włóknem mięśniowym-zwane złączem nerwowo-mięśniowym (NMJ). W tym miejscu włókno mięśniowe po raz pierwszy reaguje na sygnalizację przez neuron ruchowy. Każde włókno mięśni szkieletowych w każdym mięśniu szkieletowym jest unerwione przez neuron ruchowy w NMJ. Sygnały wzbudzenia z neuronu są jedynym sposobem na funkcjonalne aktywowanie włókna do kurczenia się.
sprzężenie pobudzająco-skurczowe
wszystkie żywe komórki mają potencjały błonowe lub gradienty elektryczne na ich błonach. Wewnątrz membrany jest zwykle około -60 do -90 mV, w stosunku do zewnątrz. Jest to określane jako potencjał błonowy komórki. Neurony i komórki mięśniowe mogą wykorzystywać swój potencjał błonowy do generowania sygnałów elektrycznych. Robią to, kontrolując ruch naładowanych cząstek, zwanych jonami, przez membrany, aby wytworzyć prądy elektryczne. Osiąga się to poprzez otwieranie i zamykanie wyspecjalizowanych białek w błonie zwanych kanałami jonowymi. Chociaż prądy generowane przez jony poruszające się przez te białka kanałów są bardzo małe, stanowią one podstawę zarówno sygnalizacji nerwowej, jak i skurczu mięśni.
zarówno neurony, jak i komórki mięśni szkieletowych są elektrycznie pobudliwe, co oznacza, że są w stanie generować potencjały działania. Potencjał czynnościowy jest specjalnym rodzajem sygnału elektrycznego, który może przemieszczać się wzdłuż błony komórkowej jako fala. Pozwala to na szybką i wierną transmisję sygnału na duże odległości.
chociaż termin sprzężenie pobudzająco-skurczowe dezorientuje lub przeraża niektórych uczniów, sprowadza się do tego: aby włókno mięśni szkieletowych mogło się kurczyć, jego błona musi być najpierw „wzbudzona”—innymi słowy, musi być stymulowana, aby wystrzelić potencjał działania. Potencjał działania włókien mięśniowych, który przemieszcza się wzdłuż sarcolemmy jako fala, jest „sprzężony” z faktycznym skurczem poprzez uwalnianie jonów wapnia (Ca++) z SR. po uwolnieniu Ca++ wchodzi w interakcję z białkami ekranującymi, zmuszając je do odsunięcia się tak, że miejsca wiążące aktynę są dostępne do przyłączenia przez głowy miozyny. Miozyna następnie ciągnie włókna aktyny w kierunku środka, skracając włókno mięśniowe.
w mięśniach szkieletowych sekwencja ta zaczyna się od sygnałów z somatycznego podziału ruchowego układu nerwowego. Innymi słowy, etap „wzbudzenia” w mięśniach szkieletowych jest zawsze wyzwalany przez sygnalizację z układu nerwowego (Rysunek 4).
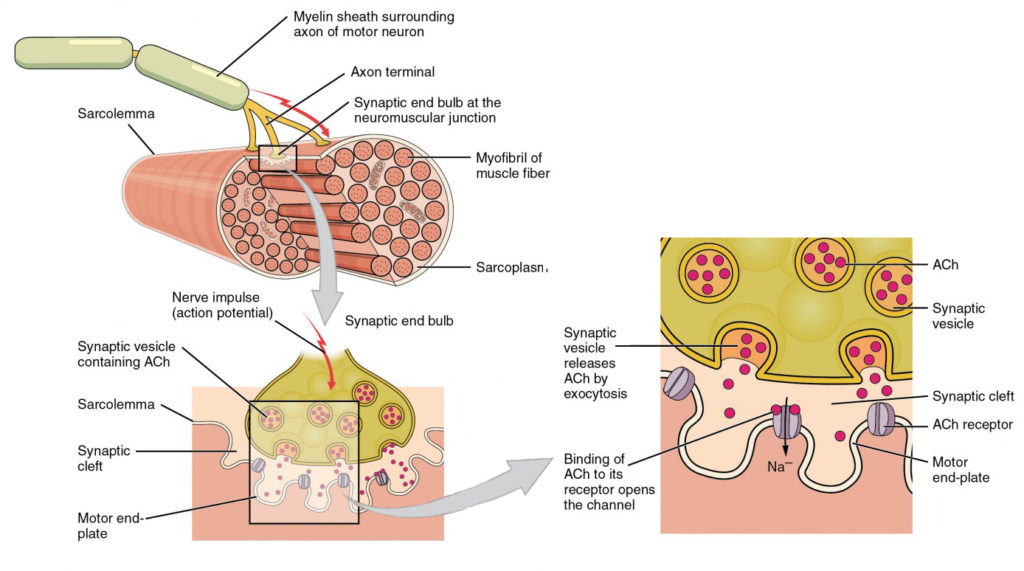
Rysunek 4. Motor End-płyta i unerwienie. W NMJ Terminal axon zwalnia ACh. Motor end-plate jest miejscem receptorów ACh w sarcolemma włókien mięśniowych. Kiedy cząsteczki ACh są uwalniane, dyfundują w przestrzeni minutowej zwanej rozszczepem synaptycznym i wiążą się z receptorami.
neurony ruchowe, które informują włókna mięśni szkieletowych o kurczeniu się, pochodzą z rdzenia kręgowego, z mniejszą liczbą zlokalizowaną w pniu mózgu w celu aktywacji mięśni szkieletowych twarzy, głowy i szyi. Neurony te mają długie procesy, zwane aksonami, które specjalizują się w przenoszeniu potencjałów działania na duże odległości— w tym przypadku, aż od rdzenia kręgowego do samego mięśnia (który może znajdować się do trzech stóp dalej). Aksony wielu neuronów łączą się ze sobą tworząc nerwy, jak przewody połączone razem w kabel.
Sygnalizacja rozpoczyna się, gdy neuronalny potencjał działania przemieszcza się wzdłuż aksonu neuronu ruchowego, a następnie wzdłuż poszczególnych gałęzi, aby zakończyć się w NMJ. W NMJ Terminal aksonu uwalnia chemiczny przekaźnik lub neuroprzekaźnik, zwany acetylocholiną (ACh). Cząsteczki ACh dyfundują w przestrzeni minutowej zwanej rozszczepem synaptycznym i wiążą się z receptorami ACh zlokalizowanymi w motorycznej płytce końcowej sarcolemmy po drugiej stronie synapsy. Gdy ACh wiąże się, kanał w receptorze ACh otwiera się i dodatnio naładowane jony mogą przejść do włókna mięśniowego, powodując jego depolaryzację, co oznacza, że potencjał błonowy włókna mięśniowego staje się mniej ujemny (bliżej zera.)
gdy membrana się depolaryzuje, uruchamiany jest kolejny zestaw kanałów jonowych zwanych napięciowymi kanałami sodowymi. Jony sodu dostają się do włókna mięśniowego, a potencjał czynnościowy szybko rozprzestrzenia się (lub „ognie”) wzdłuż całej błony, aby zainicjować połączenie wzbudzająco-skurczowe.
w świecie pobudliwych błon dzieje się bardzo szybko (pomyśl tylko, jak szybko możesz pstryknąć palcami, gdy tylko się na to zdecydujesz). Natychmiast po depolaryzacji błony repolaryzuje się, przywracając ujemny potencjał błonowy. Tymczasem ACh w rozszczepieniu synaptycznym jest rozkładany przez enzym acetylocholinoesterazę (AChE), tak że ACh nie może ponownie połączyć się z receptorem i ponownie otworzyć swojego kanału, co spowodowałoby niepożądane wydłużone pobudzenie i skurcz mięśni.

Rysunek 5. T-tubule. Wąskie rurki T umożliwiają przewodzenie impulsów elektrycznych. Funkcja SR reguluje wewnątrzkomórkowy poziom wapnia. Dwie końcowe cysternae (gdzie powiększony SR łączy się z kanalikiem T) i jeden kanalik t tworzą triadę-„trójkę” błon, z tymi z SR po dwóch stronach i kanalikiem t umieszczonym między nimi.
propagacja potencjału czynnościowego wzdłuż sarcolemmy jest częścią wzbudzenia sprzężenia wzbudzenie-skurcz. Przypomnijmy, że to wzbudzenie faktycznie wyzwala uwalnianie jonów wapnia (Ca++) z jego przechowywania w komórce SR. Aby potencjał czynnościowy dotarł do błony SR, w sarcolemmie występują okresowe inwaginacje, zwane kanalikami T („T” oznacza „poprzeczny”). Przypomnijmy, że średnica włókna mięśniowego może wynosić do 100 µm, więc te kanaliki T zapewniają, że błona może zbliżyć się do SR w sarkoplazmie. Układ rurki T z membranami SR po obu stronach nazywa się triadą (ryc. 5). Triada otacza cylindryczną strukturę zwaną miofibrilem, która zawiera aktynę i miozynę.
kanaliki t przenoszą potencjał czynnościowy do wnętrza komórki, co powoduje otwarcie kanałów wapniowych w błonie sąsiedniego SR, powodując rozproszenie Ca++ z SR do sarkoplazmy. To właśnie pojawienie się Ca++ w sarkoplazmie inicjuje skurcz włókna mięśniowego przez jego jednostki skurczowe lub sarkomery.