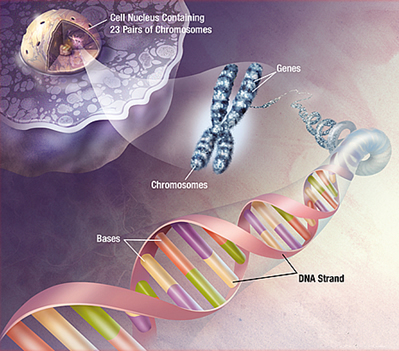
植物のDNA
DNAは、生物の構造と機能の情報を運ぶすべての細胞に存在する遺伝性ま
植物界では、DNA、またはデオキシリボ核酸は、核、ミトコンドリア、および葉緑体の膜結合細胞構造内に含まれています。 DNAは、化学分子の中でユニークないくつかの特性を持っています。
それはすべての生きている生物に普遍的であり、それぞれで同じ構造と機能を持っています。 それは自己複製として知られているプロセスで自分自身を再生することができます。 この特性は、細胞分裂、したがって連続性、成長、および修復を可能にする。


それは、その構造に遺伝コード、または細胞の開発と維持のための命令のセットを運ぶ。 最後に、それは進化、多様性、および病気に寄与する突然変異と呼ばれる環境および内部の原因の両方から、化学構造の変化を受ける。
化学構造
DNAは四つのヌクレオチドからなる単純な分子です。 各ヌクレオチドは、5炭素糖(デオキシリボース)、リン酸、および4つの可能な窒素塩基のいずれかを持っています:アデニン(A)とグアニン(G)の二重環プリンとチミン(T)とシトシン(C)の一重環ピリミジン。
DNAの特性のほとんどは、ヌクレオチド間で形成されるユニークな結合に関連しています:糖リン酸成分は直線的に整列し、窒素環は垂直に結合します。

化学構造
窒素環はさらに非常に特定の方法で結合します:常にTとペア、およびCとゴールウェイ対DNAmoleculeは、このようにラダー、側が糖-リン酸であること、ラング、A-TおよびG-C対。
さらに結合と折り畳みは、二重らせんとして知られている螺旋状のはしごのような形をした構造を生成します。 この二重らせんは、細胞分裂前および分裂中に光学顕微鏡下で見える染色体として知られるロープ状の構造にコンパクトに包装されている。
細胞の日常生活の間、DNAはクロマチン(真核細胞の核に位置するDNAとそれに結合するタンパク質を指す包括的な用語)と区別できないダークマスコールとし
ワトソンとクリックモデル
1800年代半ばのオーストリアの修道士グレゴール-メンデルは、遺伝物質が存在すると仮定した。 彼は、花の色などの形質の継承を研究するために、彼の庭のエンドウ豆や他の植物を使用して遺伝の法則を発見しました。
p>
科学者ジェームズ-ワトソンとフランシス-クリックは、1953年に、分子のユニークな特性のそれぞれのための最ももっともらしいモデルとして二重らせん それらのモデルは、その後すぐにX線回折技術によって検証された。
コロンビア大学や米国の他の場所で働いているいくつかの研究者は、ワトソンとクリックの前に、この遺伝物質の化学組成と窒素塩基対を発見することによって道を導いていた:アデニンの量は常にチミンの量とグアニンとシトシンの量と同等であった。
ワトソンとクリックモデルはまた、DNAの両側、またはストランドは、反対方向に実行することを示唆した:つまり、一方の側のリン酸糖は上向きに指し、他方のストランドは下向きに指しています。
このプロパティは反平行結合として知られています。 WatsonとCrickモデルは、細胞分裂中にDNAがどのように複製され、遺伝情報がその構造にどのようにコードされるかを簡単に説明することができます。
自己複製
世代の継続性と個々の生物の成長と修復を可能にする自己複製は、細胞分裂中に起こる。 DNAはそれ自身の正確なコピーを生成することができなければなりません。 分子はこれのために独特に設計されている:一連の酵素仲介されたステップは二重螺旋が、2本の繊維を分けるジッパーのように、ほどくか、または解
次に、消化された食物からのヌクレオチドが最初に細胞に入り、次に核に入ります。 このプロセスは、DNAの二つの新しい二本鎖分子が形成され、それぞれが他のコピーを形成し、それぞれが細胞分裂に起因する新しい細胞に入るまで続
タンパク質合成
DNAにコードされた情報は、細胞と生物のすべての発達と維持を可能にします。 このコードの言語は、各鎖上の隣接するヌクレオチドの線形読み取りにあります。 すべての三つのヌクレオチドは、特定のアミノ酸、タンパク質の個々の単位を指定または適合させます。
第二の分子、リボ核酸(RNA)は、DNAの分子構造をコピーし、アミノ酸がタンパク質を生成するために、指定された順序で、組み立てられている細胞の周囲の細胞質に核外の情報をもたらします。
糖、脂肪、または金属の添加など、これらのタンパク質の生産後の変更は、機能的および構造的多様性の広大な配列を可能にします。 植物DNAは植物に独特であるいろいろな物質のためにコードします。 これらの製品は、植物自体だけでなく、人類だけでなく生態学的なニッチ全体も維持しています。
ミトコンドリアと葉緑体DNA
第二に、独立して機能するDNAのセットは、細胞の核、ミトコンドリアと葉緑体の外側の二つのオルガネラに存在します。 それはmitochondria、炭水化物、脂肪および蛋白質が熱(カロリー)の形で貯えられた化学結合エネルギーの解放の未加工要素に破壊される細胞の動力源にある。
DNAが核の外側に収容されている第二の領域は、植物細胞に特有の構造である葉緑体にあります。 葉緑体では、光合成が起こり、植物が二酸化炭素、水、太陽エネルギーを変換して糖を生成し、後に脂肪およびタンパク質を生成し、酸素を放出するプロセス 植物によって行われたこの重要なプロセスは、地球上のほとんどの生命を維持します。
ミトコンドリアDNAと葉緑体DNAの両方が、細胞分裂中に核DNAとは別に複製する。 これらのオルガネラは、数十億年前に、真菌、原生生物、植物、動物などの非細菌生命体を構成する真核細胞を形成するために他の細胞に組み込まれた独立
植物タンパク質
植物に固有のタンパク質の大規模な配列は、植物DNA上にコードされています。 多くの注目を集めているグループは、いわゆる植物化学物質であり、強力な健康上の利点を有する物質である。 よく研究されたクラスは、フラボノイド、フィトステロール、カロテノイド、インドール、クマリン、有機硫黄、テルペン、サポニン、リグナン、イソチオシアネートを含む、いくつかである。
各グループには、抗酸化物質と抗カルシノーゲンの両方である特定のタンパク質が含まれており、動物細胞を癌原因物質から保護しています。 オレンジと黄色の果物や野菜に含まれるベータカロチン、トマトに含まれるリコピンなどのカロテノイドは、心臓病や脳卒中、癌から動物を保護するように見えます。
植物ステロールは、大豆に見られるもののように、女性ホルモンを模倣するエストロゲンのような化合物です。 これらは癌から女性器官を保護し、またコレステロールを下げるようであるようです。
植物ホルモン
植物DNAの大きなセグメントは、特殊な植物ホルモンのコーディングに専念しています。 ホルモンは、ある細胞群によって産生され、別の部位に循環し、標的細胞のDNAに影響を与える物質である。 植物では、これらのホルモンは細胞分裂、成長および微分を制御します。
植物ホルモンの五つのよく説明されたクラスがあります: オーキシン、ジベレリン、サイトカイニン、エチレン、アブシジン酸。 オーキシンの機能の中には、植物を光に向かって曲げる性質である光刺激を可能にすることがあります。
根で生産され、オーキシンは茎に移動し、細胞を植物組織の暗い側で伸長させる。 エチレンは、果実を熟させ、植物から落下させる気体物質である。 アブシジン酸は、葉の老化および落下に寄与する。
遺伝子組み換え植物
植物は操作が容易であるため、植物DNAは生物工学者のための主要な実験対象として細菌DNAに次ぐものである。 特定の形質をコードする遺伝子の特定のセグメントを追加または削除することによるDNAの直接修飾は、生物工学およびバイオテクノロジーの焦点で
植物は人間や家畜の人口のための主要な食料源を提供するため、昆虫、細菌、ウイルス、および他の害虫に抵抗し、外部農薬の必要性を減少させる遺伝子組換え食品が開発されている。
遺伝子組み換え植物作物は、見て、おいしいから速く成長したり、種子を持たないことに、よりゆっくりと熟成に、さまざまな特性を強化するように設計されています。
動物界などの他の王国の遺伝子を植物DNAに導入することで、科学者はヒトワクチン、ヒトホルモン、その他の医薬品を含む可能性のある将来の作物を開発することができるようになった。
トマトは、米国で販売される最初の連邦政府承認の生物工学食品でした。 今日では、何十もの農産物や家畜飼料が何らかの形で遺伝子組み換えされています。


